
1 INTRODUCCIÓN AL CRECIMIENTO BACTERIANO
3 MEDIDA DEL CRECIMIENTO DE POBLACIONES BACTERIANAS
3.2 MEDIDA DEL NÚMERO DE INDIVIDUOS
4 CRECIMIENTO BALANCEADO (= EQUILIBRADO)
5 CULTIVO CONTINUO (SISTEMAS ABIERTOS). QUIMIOSTATO
6 CULTIVO EN SISTEMAS CERRADOS
6.1 CURVA DE CRECIMIENTO EN UN SISTEMA CERRADO EN MEDIO LÍQUIDO
![]()
Se suele definir el crecimiento de cualquier sistema biológico como el incremento ordenado de todos los elementos componentes de ese sistema, lo cual implica un aumento de la masa celular que eventualmente conduce a la multiplicación celular. En organismos pluricelulares dicha multiplicación se traduce en un aumento del tamaño del individuo, mientras que en unicelulares que se dividen por fisión o por gemación, lo que ocurre es un aumento de la población. En los microorganismos cenocíticos (en los que la duplicación del genoma no se acompaña de divisón celular) el crecimiento se traduce en aumento de tamaño de la “colonia” cenocítica. El crecimiento bacteriano podemos estudiarlo desde dos puntos de vista:
|
A escala individual | |
|
A escala poblacional |
Los eventos de crecimiento en el ámbito individual hacen referencia al ciclo celular, y dentro de éste podemos abordar los siguientes temas:
|
inicio y transcurso de la replicación cromosómica y de plásmidos (véase tema 7); | |
|
segregación de cromosoma y plásmidos a las células hijas; | |
|
síntesis de nuevos materiales de las envueltas, sobre todo de pared celular (tema 5); | |
|
señales que coordinan la replicación genómica con la división celular. |
El estudio del crecimiento a nivel de poblaciones incluye los siguientes tópicos:
|
cinética del crecimiento; | |
|
factores que afectan al tiempo de generación (g); | |
|
factores ambientales que limitan el crecimiento. |
En el presente capítulo nos dedicaremos al estudio de algunos aspectos del ciclo celular procariótico, pero sobre todo nos centraremos en el crecimiento de poblaciones (que es el más frecuentemente empleado al trabajar con microorganismos), con una visión de algunos métodos para obtener crecimientos balanceados. La influencia de los factores ambientales sobre el crecimiento son el objeto de los temas 13 (agentes físicos) y 14 (agentes químicos).
Un ciclo celular es una secuencia de acontecimientos interconectados que comienza con la formación de una nueva célula y termina cuando dicha célula se divide en otras dos hijas. Los ciclos celulares se describen generalmente atendiendo a una serie de eventos identificables que se van produciendo en una secuencia fija, de modo que para que se produzca cada uno de ellos hace falta que se complete el anterior. Los periodos del ciclo celular procariótico son:
|
fase C (equivalente a la S eucariótica): replicación del ADN cromosómico; | |
|
fase D (equivalente a G2 + M): se distingue porque su terminación coincide con el final de división celular; | |
|
fase innominada, equivalente a la G1 eucariótica. |
Lo característico del ciclo es que las fases C y D son relativamente constantes, y por lo tanto, cuando disminuye el tiempo de generación (g), lo hace a expensas de la fase “G1”. Por ejemplo, en Escherichia coli, C = unos 40 min, y D = unos 20 min.
Vamos a estudiar con el ejemplo de E. coli qué ocurre con el ciclo celular a distintos tiempos de generación.
|
Cuando g>C+D |
Existe fase "G1" (intervalo que precede a la replicación). En estas condiciones podemos observar nítidamente periodos diferenciados entre sí (de síntesis de ADN y de división celular). |
|
Cuando g=C+D |
Ya no existe G1, y cada ronda de replicación comienza inmediatamente tras la precedente división celular (es decir, cada célula hija recién nacida se embarca inmediatamente en replicar el cromosoma, y una vez que ha terminado de replicarlo, la célula inicia la división celular). |
|
Cuando g<C+D |
Las rondas de replicación de un ciclo se inician antes de que haya terminado la división celular del anterior (superposición parcial entre fases C y D). |
|
Cuando g<C |
ya no se puede detectar fase D como tal. La síntesis de ADN es continua durante todo el ciclo. Se inician rondas de replicación cromosómica antes de que haya terminado la anterior. Esto se traduce en que los cromosomas muestran un típico patrón de replicación dicotómica y en que dichos cromosomas pasan a cada célula hija ya embarcados en una nueva ronda de replicación. |
Una característica del ciclo es que, cuanto más rico es un medio, y por lo tanto menor es el tiempo de generación (g), mayor es el tamaño medio de las células. Es decir, los individuos de una cepa bacteriana que esté en un medio rico son más grandes que los individuos de esa misma cepa creciendo en un medio pobre.
De lo anterior se deduce que debe de existir algún tipo de acoplamiento entre las ondas de replicación y la división celular. Este es un campo de investigación aún joven, del que no conocemos todavía todos los detalles.
Las copias de cromosomas recién replicadas se encuentran en principio adyacentes de la membrana citoplásmica (¿unidas a sendos mesosomas?) probablemente unidas a través de sus respectivos orígenes de replicación (oriC). No se sabe muy bien cómo esas copias se van separando de modo que cada una va a parar a una célula hija. Se sabe que en este reparto o partición de los cromosomas intervienen varias proteínas, entre ellas ParA y ParB, que en las células a punto de dividirse se sitúan en los dos polos opuestos. Es posible que exista algún mecanismo “activo” para separar estos cromosomas, pero parece ser que también intervienen los mecanismos “pasivos” de crecimiento de membrana y pared celular en el tabique transversal en crecimiento.
También se sabe hoy que existen dos secuencias paralelas e independientes de acontecimientos que controlan el ciclo celular: un control es sensible a una masa umbral celular para desencadenar el inicio de la replicación del cromosoma, y el otro responde a cierta longitud umbral (en el caso de los bacilos) para que ocurra el reparto de los cromosomas y la formación del tabique.
|
El comienzo de la replicación del cromosoma se indica mediante la unión de muchas unidades de la proteína DnaA (unida a ATP) al origen de replicación (oriC). Una vez que se ha iniciado una ronda de replicación en oriC, esta región queda hemimetilada, pero en vez de ser metilada inmediatamente, queda secuestrada u ocultada a efectos de la metilasa Dam y de la maquinaria de replicación. Se sugiere que en este fenómeno está implicada una proteína (SeqA), que a su vez ayudaría a esconder a oriC en la membrana. Sólo más tarde (y por factores desconocidos, pero que puede que tengan que ver de nuevo con la proporción entre sitios de inicio y algún parámetro celular como masa o volumen), vuelven a quedar las secuencias oriC disponibles para su metalición y uso en una nueva ronda. |
NOTA: La metilasa Dam es una enzima que metila las dos adeninas de la secuencia GATC/CTAG. El alumno seguramente sabrá ya que esta metilasa metila las adeninas de esa secuencia a partir de ADN hemimetilado. El ADN de la célula está totalmente metilado en esas secuencias, pero cuando pasa la horquilla de replicación, la cadena recién sintetizada carece de esa modificación, hasta que llega la metilasa Dam, que reconoce la secuencia hemimetilada y metila la adenina de la cadena no metilada. Un rasgo curioso de la región oriC para el inicio de la replicación cromosómica es que contiene una proporción de secuencias GATC/CTAG muy superior a la media del resto del genoma.
|
El comienzo de la tabicación parece que requiere dos señales:
|
Como ya vimos en el capítulo 5, llegado el momento, la proteína FtsZ (que hasta entonces estaba diseminada por toda la célula), se concentra ahora en la parte central, señalando el plano de la tabicación: se forma un “anillo citocinético” o divisoma, en el que la FtsZ se va “contrayendo” hacia el interior (hidrolizando GTP hasta GDP). A continuación se van ensamblando en el divisoma varias proteínas Fts, entre ellas la PBP3, que como vimos es una transglucosidasa-transpeptidasa específica del tabique, que va produciendo peptidoglucano en el septo transversal. La terminación del tabique señala el final del ciclo celular.
En la práctica habitual del laboratorio, los experimentos se realizan siempre con poblaciones bacterianas, a menudo tomando muestras en diversos momentos de su crecimiento en un medio de cultivo. En el resto del capítulo nos vamos a referir al crecimiento microbiano a escala de poblaciones. Comenzaremos con algunos métodos habituales de medir ese crecimiento poblacional, algunos de los cuales serán aprendidos “en vivo” por el alumno en las prácticas de laboratorio.
El crecimiento de una población o cultivo bacterianos se puede expresar en función de:
|
aumento de masa del cultivo | |
|
aumento del número de células |
Ambos tipos de expresiones son equivalentes entre sí en cultivos que estén en crecimiento balanceado o equilibrado (en el que todos los componentes aumentan una misma proporción por unidad de tiempo).
En estos métodos se requieren preparaciones limpias, sin partículas extrañas.
1) Determinación del peso húmedo:
|
se tara un tubo de centrífuga; | |
|
se centrifuga el cultivo y se elimina el sobrenadante; | |
|
se determina el peso del sedimento. |
Inconvenientes: grandes errores, debido al líquido intercelular retenido, cuya cuantía depende a su vez de la forma y tipo de agrupaciones de la cepa, intensidad del empaquetamiento, etc.
2) Determinación del peso seco: como el anterior, pero el sedimento se seca antes de ser pesado (105ºC, toda la noche), hasta peso constante. Las medidas de peso seco suelen representar el 10-15% de los valores de peso húmedo.
Inconvenientes: método tedioso (requiere mucho tiempo) y con bastantes errores: es difícil pesar menos de 1 mg con exactitud en las balanzas habituales de laboratorio. 1 mg de peso seco equivale a unas 5x109 bacterias.
3) Determinación del nitrógeno total: técnica de micro-Kjeldahl.
4) Determinación de un componente característico: peptidoglucano, ADN, ARN, proteínas, ATP, clorofilas en organismos fotosintéticos, etc.
Comentarios: se suele usar en bacterias para las que otros métodos más fáciles dan errores debido a que forman grumos no dispersables, crecen en filamentos, etc. Se emplean en determinaciones de crecimientos en ambientes naturales.
1) Medida de consumo de nutrientes o de producción de algún metabolito por unidad de tiempo . Ejemplos: consumo de oxígeno (QO2) y consumo de carbónico (QCO2), determinados por el respirómetro de Warburg. Producción de ácidos.
2) Métodos turbidimétricos (ópticos). Son muy usados en la práctica cotidiana del laboratorio. La base común de estos métodos consiste en la medición de la cantidad de luz dispersada o transmitida a través de un cultivo bacteriano.
Recordemos aquí que las suspensiones bacterianas dispersan la luz, al igual que cualquier partícula pequeña suspendida en agua (efecto Tyndall). La dispersión de la luz es, dentro de ciertos límites, proporcional a la masa del cultivo.
a) Espectrofotómetro: Este aparato es de uso habitual en cualquier laboratorio de Microbiología o Bioquímica. Mide la densidad óptica (D.O.), es decir la absorbancia (una medida de la luz transmitida a través de la supensión).
Por supuesto, hay que realizar una curva estándar para relacionar los valores de A con la masa bacteriana en una muestra problema.
Comentarios: La cantidad de luz dispersada es proporcional al cociente entre el tamaño de la partícula y la longitud de onda incidente; la sensibilidad de la técnica aumenta pues a longitudes de onda (l) cortas. La proporcionalidad entre A y masa bacteriana sólo es válida para >107céls/ml.
b) Nefelómetro: Es un aparato similar al anterior, pero el dispositivo sensor está situado en ángulo recto respecto de la dirección de la luz incidente, y lo que se mide es pues, la luz dispersada directamente por la preparación. Posee mayor sensibilidad que el espectrofotómetro.
1) Cámara de recuento de Petroff-Hauser: Consiste en un portaobjetos especial con una graduación en superficie y unas medidas muy concretas:
|
excavación con 0.02 mm de profundidad; | |
|
área de 1 mm2, dividida en un retículo de 25 cuadrados grandes; | |
|
cada cuadrado grande está subdividido a su vez en 4x4 = 16 cuadrados pequeños. | |
|
O sea, la muestra se distribuye en 16 x 25 = 400 celdillas (cuadros pequeños). |
La muestra, una vez dispensada entre el porta y el cubre, se deja reposar sobre la plataforma del microscopio durante unos minutos, y se cuenta el número de células en varias celdillas (normalmente en 16, equivalentes a uno de los cuadros grandes). Se anota el número n de células observadas en esas 16 celdillas. Entonces, la concentración celular es fácil de establecer:
n x 25 x 50 x 1000 = concentración en células/ml.
Ventajas: es un método muy rápido.
Inconvenientes: sólo sirve para suspensiones relativamente concentradas (>10x106 céls./ml). Por debajo de este valor el número de células vistas en el campo del microscopio es muy pequeño y poco significativo estadísticamente. En bacterias móviles, hay que inmovilizarlas previamente, con una mezcla de alcohol y agua.
2) Contadores electrónicos de partículas (tipo Coulter): Se hace pasar una suspensión microbiana por un tubo capilar, entre los dos polos de una corriente eléctrica. Cada vez que por un orificio (30 mm diámetro) pasa una partícula (p. ej., bacteria) se interrumpe la corriente, lo cual es recogido por un dispositivo de registro electrónico, que detecta el número y el tamaño de las partículas que van pasando. (El tamaño detectado es función de la intensidad del pulso de voltaje al paso de la partícula).
Comentarios: hay que usar suspensiones absolutamente libres de partículas extrañas (las pequeñas serían contabilizadas erróneamente como bacterias, y las mayores pueden obturar el orificio del aparato).
1) Recuento de viables en placa: Los métodos de recuento de número de células que hemos visto hasta ahora (los directos) no distinguen entre células vivas y muertas. En muchos casos conviene contar las células vivas, y esto en laboratorio se suele hacer mediante el recuento de viables. (Una célula se define como viable cuando, colacada en un medio adecuado, es capaz de dividirse y dar descendencia). El método habitual de lograr esto es sembrar pequeñas alícuotas de diluciones adecuadas de un cultivo original sobre placas de Petri con medio sólido (con agar). Cada célula viable dará origen a una colonia visible después del tiempo adecuado de incubación. Contando las colonias visibles, teniendo en cuenta la dilución de la que proceden y el volumen de alícuota utilizado, es fácil deducir el número de células viables en la suspensión original. (Para esto, mira el ejemplo de la diapositiva, y sobre todo, estate atento a la práctica correspondiente, que se suele realizar en la 3ª tanda).
Mientras tanto, y para luego repasar esa práctica, puedes realizar un experimento on-line sobre crecimiento bacteriano
Precauciones:
|
para minimizar los errores estadísticos de muestreo, se recomienda sembrar 5 placas de cada dilución; | |
|
hay que usar pipetas nuevas en cada dilución; | |
|
contar las placas donde existan entre 50 y 300 colonias. | |
|
Como no podemos garantizar que cada colonia no proceda de más de un indiviudo (y esto es especialmente cierto para bacterias que forman agrupaciones de 2 o más células), el recuento se refiere no a “células viables reales” sino a “unidades formadoras de colonia” (UFC). Por lo tanto, una UFC corresponde, como mínimo, a una bacteria, pero sobre todo en bacterias con agrupaciones, la medida por siembr en placa infravalora el número real de individuos, porque cada UFC puede corresponder a dos o más individuos que estaban juntos al ser sembrados en la placa. |
2) Recuento sobre filtros: Se usa para suspensiones diluidas de bacterias. Se hace pasar un gran volumen de suspensión a través de una membrana de nitrocelulosa o de nylon estériles (con un diámetro de poro que retiene las bacterias pero permite el tránsito de sustancias). Posteriormente, el filtro se deposita sobre la superficie de un medio de cultivo sólido. Las colonias se forman sobre el filtro a partir de las células retenidas. Dichas colonias se cuentan, deduciéndose la concentración original de viables en función del volumen de suspensión que se hizo pasar por el filtro.
Una población de bacterias que se encuentre en un medio adecuado en el que se mantienen constantes todos sus parámetros nutricionales y ambientales, crece de forma tal que el incremento por unidad de tiempo de masa celular, no de células, ADN, ARN, proteínas, etc., es un valor constante y similar en cada caso:
DM/M = DN/N = D[ADN]/[ADN] = D[proteínas]/[proteínas] = ... = K
Así pues, durante este crecimiento, de tipo exponencial o logarítmico, el cultivo se comporta como una reacción autocatalítica de primer orden:
velocidad de aumento del componente = K·{cantidad del componente}
También se puede decir que el no de células, la masa celular u otros componentes se duplican en un mismo lapso de tiempo determinado.
Este tipo de crecimiento se denomina balanceado o equilibrado. Se caracteriza, pues, por ser el crecimiento en el que
|
todos los constituyentes celulares se duplican en un mismo tiempo, o dicho de otra manera: | |
|
aquel en el que estos constituyentes aumentan proporcionalmente por un mismo factor en la unidad de tiempo. Este factor es el coeficiente exponencial de crecimiento (m), que es característico para cada cepa bacteriana en cada medio determinado. |
Para deducirla vamos a aprovechar la definición empírica anterior, atendiendo por un lado al aumento de la masa celular, y por otro al incremento del número de individuos.
a) En función del aumento de masa celular:
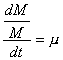 , y por lo tanto, dM = M·m·dt
, y por lo tanto, dM = M·m·dt
Si integramos, resulta: M/M0 = em(t-t0)
Aplicando logaritmos neperianos:
![]() {12.1}
{12.1}
De aquí se puede deducir que el coeficiente m es
 {12.2}
{12.2}
b) En función del aumento del número de células. Supongamos que partimos de una célula. Tras una división (generación celular), tendremos 2, tras dos divisiones tendremos 4, luego 8, etc: tenemos una serie geométrica. 1, 2, 22, 23, 24, ... 2n (donde n es el número de generaciones transcurridas). Si en vez de partir de una célula partimos de N0 células iniciales, tenemos:
N = N0·2n ; por lo tanto:
N/N0 = 2n {12.3}
Por otro lado, el número de generaciones se puede calcular fácilmente teniendo en cuenta el tiempo transcurrido y conociendo el tiempo medio de generación (g):
![]()
Sustituyendo esta expresión en la fórmula {12.3} tenemos:
N/N0 = 2(t-t0)/g
Apliquemos logaritmos neperianos:
![]() {12.4}
{12.4}
Ahora bien, como estamos ante un crecimiento balanceado, las dos expresiones matemáticas {12.1} y {12.4} que hemos deducido (la de la sección A, basada en masa, y la de la sección B, basada en número de individuos) son equivalentes:
![]() , y por lo tanto:
, y por lo tanto:
m (t-t0) = (t-t0)/g·ln2, de donde se deduce el valor del coeficiente de crecimiento exponencial:
![]() , expresado en h-1
{12.5}
, expresado en h-1
{12.5}
Esta es la expresión matemática del coeficiente del crecimiento balanceado, en función del tiempo medio de generación (g).
En un medio ideal, sin limitación de nutrientes, este coeficiente es m = mmáx, o sea, el máximo valor posible del coeficiente para esa cepa en ese medio. Es decir, es un crecimiento balanceado no restringido.
En un medio donde exista algún nutriente limitante (o sea, cuya concentración está por debajo del límite de eficiencia de las permeasas), el crecimiento balanceado es de tipo restringido, de modo que el coeficiente real de crecimiento es inferior al máximo, según la fórmula empírica siguiente (ecuación de Monod):
![]()
donde [S] es la concentración del nutriente limitante; KS es la constante de saturación, equivalente a la concentración de nutriente para la que el coeficiente es semimáximo. Como se puede ver, la tasa de crecimiento (medida por m) es una función hiperbólica de la concentración del sustratro (nutriente) limitante (ver gráfico). Este sustrato puede ser una fuente de C y/o de energía, fuente de N, de P, un factor de crecimiento, etc.
Ejemplos de KS:
|
la KS para la glucosa en E. coli es 1·10-6M | |
|
la KS para el triptófano en esta bacteria es 2·10--7 M |
Estos bajos valores se deben a la alta afinidad de las permeasas de membrana hacia los sustratos, lo cual es una adaptación evolutivamente adquirida de las bacterias a los medios muy diluidos en nutrientes en los que normalmente viven. (Por el contrario, los medios de laboratorio donde se suelen cultivar las bacterias suelen ser más concentrados).
Como veremos en el apartado 6, en la naturaleza, y en los sistemas de cultivo cerrados que habitualmente se emplean en laboratorio, tarde o temprano el crecimiento balanceado suele terminar, debido a que se agotan los nutrientes o se acumulan sustancias de desecho.
El cultivo continuo es un cultivo balanceado mantenido por tiempo indefinido por un sistema abierto de flujo que se compone de:
|
una cámara de cultivo de volumen constante, | |
|
a la que llega un suministro de nutrientes, | |
|
y de la que se eliminan o separan los productos tóxicos de desecho (por un dispositivo de rebosadero). |
Una vez que el sistema alcanza el equilibrio, el número de células y la concentración de nutrientes en la cámara permanecen constantes, y entonces se dice que el sistema está en estado estacionario, con las células creciendo exponencialmente.
Los parámetros a tener en cuenta son:
|
flujo (f), medido en ml/h | |
|
volumen de la cámara de cultivo (v, en ml) | |
|
densidad celular en la cámara (x) | |
|
factor de dilución D = f/v (en h-1). |
Existe una pérdida de células por el rebosadero: -dx/dt = x·D
El crecimiento bruto es dx/dt = x·m
Por lo tanto, el crecimiento neto es dx/dt = x·m - x·D = x·(m - D)
Si logramos que el coeficiente de crecimiento (m) se haga igual al factor de dilución (D), entonces dx/dt = 0, y por lo tanto la concentración de células se hace constante (x= x). El cultivo se encuentra entonces en estado dinámico de equilibrio. Las pérdidas de células por drenaje se compensan con las ganancias por crecimiento.
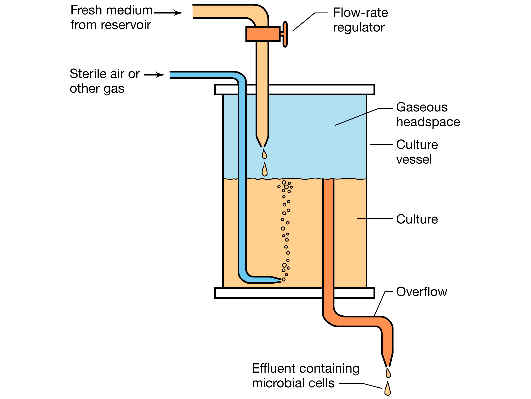
Una de las maneras de lograr un cultivo continuo es mediante el llamado quimiostato: en el quimiostato podemos controlar de modo independiente la densidad de la población celular y la velocidad de crecimiento del cultivo. La densidad celular en el equilibrio se controla ajustando el factor de dilución (D), mientras que la velocidad de crecimiento se controla variando la concentración del nutriente limitante en la cámara reservorio (SR).
|
En un quimiostato, los microorganismos pueden cultivarse a una amplia variedad de tasas de crecimiento exponencial | |
|
El quimiostato permite crecimientos balanceados restringidos debido a que existe un nutriente o sustrato presente en una concentración suficientemente baja como para limitar la densidad de población. | |
|
Así pues, el quimiostato también permite elegir la densidad de células a la que se quiere trabajar. |
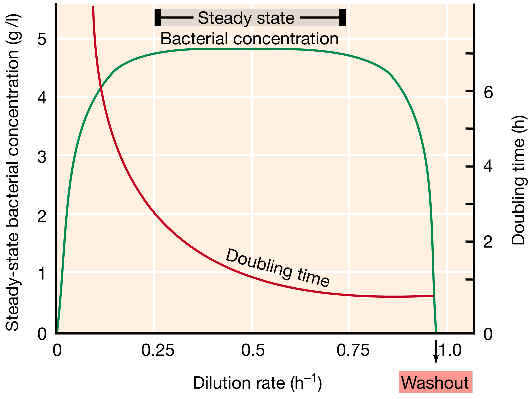
Algunos comentarios sobre el gráfico:
|
La densidad del cultivo es muy similar en un amplio margen de tasas de dilución (D). Este margen es el más adecuado para hacer estudios en el quimiostato. En cambio, el valor de tiempo de generación (g) varía ampliamente. O sea, el quimiostato puede obtener tasas de crecimiento muy diferentes, sin que se afecte la densidad celular.Sin embargo, a valores extremos de dilución, se puede ver que el equilibrio se rompe: | |
|
A altas tasas de dilución la concentración microbiana cambia rápidamente, y en un margen estrecho el cultivo puede drenarse totalmente (DC: dilución crítica). Es decir, el cultivo se “lava” porque su velocidad de crecimiento es inferior a la tasa de dilución. | |
|
A muy bajas diluciones (DM) el quimiostato no funciona si el nutriente limitante es la fuente de energía. Ello se debe a que en esas condiciones, la fuente de energía sólo se usa para reacciones de mantenimiento de la integridad celular, pero no queda para el crecimiento. A este valor lo llamamos energía de mantenimiento. Los procesos relacionados con esta energía de mantenimiento son el potencial de membrana, el transporte de solutos y la renovación de proteínas. |
Aplicaciones del cultivo continuo en quimiostato:
|
Aportan una fuente continua de células en fase exponencial, lo que se aplica a procesos industriales de fermentación (producción de bebidas alcohólicas, de antibióticos, de aminoácidos, etc). | |||||||
|
En el quimiostato, el crecimiento a bajas concentraciones de sustrato permite estudiar:
|
En los sistemas cerrados (que pueden ser líquidos o sólidos), no existe aporte continuo de nutrientes, ni drenaje de células ni de sustancias de desecho.
En estos sistemas la fase exponencial de crecimiento balanceado no restringido dura sólo unas cuantas generaciones, debido al agotamiento de nutrientes y/o a la acumulación de desechos.
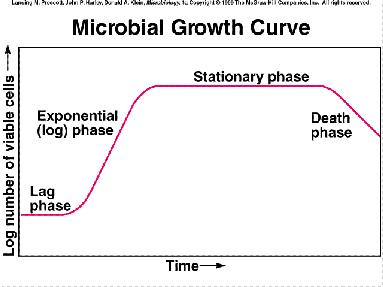
Como se puede constatar en el anterior gráfico, el crecimiento en un sistema cerrado consta de varias fases, que pasamos a comentar:
1) Fase de retardo (fase “lag”): Es el período de tiempo durante el que el inóculo se adapta a las condiciones del medio fresco sobre el que se ha sembrado. Se trata de un período de ajuste metabólico. Su duración depende de varios factores:
|
tamaño del inóculo; | |||||||
|
bondad del inóculo (estado metabólico previo del inóculo):
| |||||||
|
medio del que procede el inóculo:
|
Pero aun cuando la inoculación se hace desde un cultivo previo en fase logarítmica, cuyo medio sea idéntico al medio fresco, se observa siempre una fase lag. ¿Por qué?:
|
necesidad de neutralizar sustancias tóxicas en el medio fresco; | |
|
porque se produce dilución de ciertos metabolitos intracelulares al inocular las bacterias en el medio nuevo; por lo tanto, hasta que no se vuelva a alcanzar una concentración de esos metabolitos adecuada para el crecimiento, éste no “arranca”. |
Ejemplo: Supongamos que inoculamos una bacteria heterotrófica en un medio ligeramente ácido, sometido a aireación: en un principio, se produce dilución de CO2, por lo que se retardan reacciones de carboxilación que requieren este CO2, y se produce un retardo.
2) Fase de transición, de crecimiento acelerado, que conduce a …
3) Fase de crecimiento exponencial (= fase logarítmica). La fase 2 se debe a que cada célula entra en la fase exponencial con desfase respecto de sus compañeras. Ello demuestra que las células del inóculo no están todas en las mismas condiciones fisiológicas.
Durante la fase logarítmica se da un crecimiento balanceado no restringido durante unas pocas generaciones (normalmente menos de 10). El tiempo de generación (g) es característico para cada especie o cepa, en cada medio concreto:
El valor del tiempo de generación (g) depende de:
|
composición del medio | |
|
temperatura | |
|
pH | |
|
osmolaridad (tonicidad), etc. |
Los microorganismos heterotrofos suelen crecer más rápidamente en los medios complejos, ricos, que en los medios sintéticos, y dentro de estos últimos, mejor con glucosa que con otras fuentes de carbono. Algunos microoorganismos tienen, a su temperatura óptima tiempos de generación muy cortos (15, 20 min), mientras que otros tienen crecen más lentamente, con tiempos de generación que pueden ser de varias horas o incluso días.
4) Fase de aceleración negativa, de crecimiento desequilibrado, que conduce a…
5) Fase estacionaria: Esta fase se caracteriza porque el coeficiente neto de crecimiento se hace nulo (m = 0), pero aún existe crecimiento. Lo que ocurre es que el crecimiento bruto se equilibra con las muertes celulares.
En este período se agotan nutrientes especiales y se acumulan sustancias de desecho. Incluso el pH del medio empieza a hacerse inadecuado para el crecimiento celular.
Si la bacteria crece en un medio complejo, la fase 4 de transición (de aceleración negativa) puede ser relativamente larga, debido a que va recurriendo a fuentes alternativas (p. ej., puede recurrir a aminoácidos como fuente de C una vez agotados los hidratos de carbono).
En la fase estacionaria aún existen reacciones metabólicas, pero el metabolismo general es diferente al de la fase logarítmica:
|
las células son más pequeñas, debido a que existe división celular después de que se haya detenido el incremento de masa; | |
|
suelen ser más resistentes a agentes físicos y químicos; | |
|
existe reciclado de ciertos materiales intracelulares; | |
|
baja el contenido en ARN. |
6) Fase de transición hacia …
7) Fase de muerte exponencial: Se da muerte y lisis masiva, exponencial, del cultivo. Se debe a agotamiento de reservas de energía. Algunas veces las células aparecen grandes, hinchadas, distorsionadas (formas “fantasmas”, “ghost”). La pendiente de esta parte de la curva depende de las especies (por ejemplo, en bacterias entéricas es suave, mientras que en Bacillus es más acentuada).
Te recuerdo que puedes hacer un experimento "virtual" con la curva de crecimiento de una bacteria. Que disfrutes.
Un medio sólido es una solución nutritiva (como el líquido), pero incorporado a un gel, que le da consistencia.
Los tipos de gelificantes usados para los medios sólidos (más explicaciones en la 2ª tanda de prácticas):
|
agar-agar (o simplemente, agar): es el más comúnmente empleado; | |
|
gelatina (inconveniente de que se licúa a temperaturas relativamente bajas, y además, algunos microorganismos lo degradan por gelatinasas); | |
|
silicagel (o gel de sílice): tedioso de preparar. Uso casi exclusivo para quimioautotrofos. |
Los medios sólidos se suelen inocular mediante asa de siembra o espátula, diseminando las bacterias sobre su superficie libre, en recipientes adecuados, como las placas de Petri (ver prácticas). Tras la incubación a la temperatura y condiciones pertinentes, cada bacteria o agrupación bacteriana que ha quedado en un punto determinado del medio da origen, por crecimiento, a una acumulación de células, visible a simple vista, denominada colonia.
La densidad de cada colonia es muy alta (del orden de 107 células para una colonia de unos 5 mm). Esto se debe a que en el medio sólido, a diferencia del líquido, las bacterias no pueden dispersarse, y durante mucho tiempo este medio sólido permite un aporte continuo de nutrientes (por difusión desde el entorno de la colonia, hacia ella), y eliminación continua de productos de desecho (por difusión desde la colonia hacia fuera). Por lo tanto, se parece a un cultivo continuo, excepto que no hay drenaje de células.
Como el alumno comprobará en prácticas (2ª tanda), cada especie bacteriana suele originar colonias de un tipo determinado, en cada medio concreto. Con vistas a la determinación taxonómica, se suele tomar nota de una serie de características de las colonias (caracteres culturales):
|
tamaño (relativo) | |
|
forma general | |
|
forma de los bordes de la colonia | |
|
aspecto de la superficie y elevación sobre el sustrato | |
|
color | |
|
consistencia, etc. |
![]()



