
1 CITOPLASMA BACTERIANO: VISIÓN DE CONJUNTO
2 MATERIAL GENÉTICO: EL NUCLEOIDE
2.1.1 MÉTODOS DE OBSERVACIÓN Y ESTUDIO
2.1.2 COMPOSICIÓN QUÍMICA, ESTRUCTURA Y ORGANIZACIÓN DEL CROMOSOMA
2.2.1 DEFINICIÓN Y CONCEPTOS GENERALES
2.2.2 FENOTIPOS DETERMINADOS POR LOS PLÁSMIDOS
2.2.3 REPLICACIÓN DE LOS PLÁSMIDOS
2.2.4 REGULACIÓN DEL NÚMERO DE COPIAS
2.2.5 REPARTO DE LAS COPIAS A LAS CÉLULAS HIJAS
2.2.6 INCOMPATIBILIDAD ENTRE PLÁSMIDOS Y GRUPOS DE INCOMPATIBILIDAD
2.3 ELEMENTOS GENÉTICOS MÓVILES
3 RIBOSOMAS; SÍNTESIS Y PLEGAMIENTO DE PROTEÍNAS
3.1 ESTRUCTURA DEL RIBOSOMA EUBACTERIANO
3.2 EL MECANISMO DE LA SÍNTESIS DE PROTEÍNAS
3.3 EL PLEGAMIENTO DE LAS PROTEÍNAS: PAPEL DE LAS PROTEÍNAS CELADORAS (“CHAPERONAS”)
4 INCLUSIONES Y ORGÁNULOS PROCARIOTAS
4.1.1 INCLUSIONES POLISACARÍDICAS
4.1.2 GRÁNULOS DE POLI-ß-HIDROXIBUTÍRICO (PHB) Y DE POLI-HIDROXIALCANOATOS
4.1.3 INCLUSIONES DE HIDROCARBUROS
4.3 ORGÁNULOS PROCARIÓTICOS CITOPLÁSMICOS
![]()
El citoplasma bacteriano es la masa de materia viva delimitada por la membrana citoplásmica. En su interior se albergan:
|
cuerpos nucleares (nucleoide); | |
|
plásmidos (no en todas las cepas bacterianas); | |
|
ribosomas; | |
|
inclusiones (no en todas); | |
|
orgánulos (no en todas). |
Al igual que en los demás seres vivos, el citoplasma es un sistema coloidal cuya fase dispersante es agua junto con diversas sustancias en solución (citosol), y cuya fase dispersa está constituida por macromoléculas y conjuntos supramoleculares (partículas submicroscópicas). La viscosidad es mayor que la del citoplasma eucariótico, estando desprovisto de corrientes citoplásmicas.
Observación:
A microscopía óptica, obviamente es poco lo que se puede distinguir en él. En las células jóvenes se suele teñir de modo uniforme, teniendo un carácter basófilo (debido a la abundancia de ARN). En las células viejas se tiñe irregularmente, debido a la aparición de inclusiones y a la acumulación de sustancias de desecho.
A microscopía electrónica destaca el carácter granulado, producido por los numerosos ribosomas (que a los aumentos habituales aparecen como partículas esféricas), aunque se observa una zona irregular hacia el centro, más transparente a los electrones, que se debe a los cuerpos nucleares (nucleoide). En los intersticios entre las partículas granuladas existe una sustancia amorfa en la que no se pueden distinguir más detalles, y que corresponde a la fase dispersante acuosa de la que hablábamos más arriba.
La nueva visión del citoplasma bacteriano (2005)
Los nuevos métodos citológicos y moleculares nos están dando una nueva visión en la que el citoplasma bacteriano está más organizado y es más dinámico de lo que pensábamos hasta hace poco. Resumiremos esta nueva imagen:
|
en el citoplasma bacteriano hay una clara separación espacial y funcional entre la zona central, donde se localiza el nucleoide y en la que se da la transcripción, respecto de la zona periférica rica en ribosomas, en la que se da la síntesis de proteínas. | |
|
La maquinaria para la replicación del cromosoma (replisoma) se localiza en un sitio concreto, hacia el centro de la célula, y muy probablemente ligado a la membrana. | |
|
Por el replisoma fijo va pasando el cromosoma para su replicación, empezando por el origen oriC. Los cromosomas hijos van siendo desplazados hacia los polos (segregación), de modo que conforme avanza la replicación, cada oriC se sitúa cada vez más cerca de un polo celular. | |
|
La proteína FtsZ es homóloga de la tubulina eucariótica, y al comienzo de la división se polimeriza formando un anillo por debajo de la membrana en el centro de la célula. Este anillo parece servir de andamio para el “divisoma”, implicado en la formación del septo transversal que conduce al nacimiento de las dos células hijas. | |
|
Hay una proteína homóloga de la actina, que se polimeriza formando amplias hélices que recorren la célula de polo a polo, y que junto con el peptidoglucano, determina la forma celular | |
|
Se han descubierto proteínas que oscilan rápida y continuamente de un extremo a otro de la célula (en cuestión de segundos), y que parecen implicadas en la regulación del ciclo celular. |
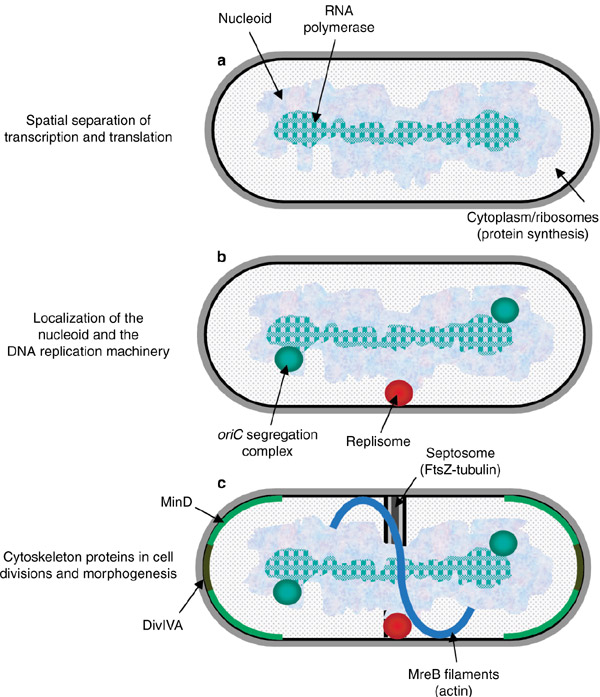
En resumidas cuentas, hoy sabemos que el citoplasma procariótico es más dinámico, estructurado y complejo que lo que creíamos hace unos pocos años.
-------------------------------
En este capítulo nos dedicaremos al estudio de las principales estructuras y macromoléculas que alberga el citoplasma. Comenzaremos con la organización a gran escala del material genético bacteriano, seguiremos con una rápida revisión sobre los ribosomas y la síntesis y plegamiento de las proteínas, y terminaremos con las inclusiones y orgánulos especiales que presentan algunas bacterias.
El ADN es el material genético de los procariotas, al igual que del resto de seres vivos (celulares). Dicho ADN está contenido en una región concreta del citoplasma, denominada nucleoide, en la mayoría de los casos sin estar separado por membrana (hay un par de bacterias en las que se ha descubierto una membrana rodeando al nucleoide). El genoma es el conjunto de genes y secuencias de ADN de un organismo. En el caso de bacterias, el elemento obligatorio del genoma es el cromosoma, aunque es frecuente encontrar unidades de replicación autónomas llamadas plásmidos, que son dispensables (si se pierden, la bacteria sigue siendo viable).
A microscopía óptica
En las células en fresco, los nucleoides se pueden poner de manifiesto (usando microscopio de contraste de fases) ajustando el índice de refracción del medio (haciendo que dicho índice sea igual al del resto del protoplasma). Se logra más detalle usando los recientes microscopios confocales de barrido.
Usando tinciones (en preparaciones fijadas previamente): se pueden emplear colorantes básicos (como los de tipo Feulgen), siempre que previamente se destruya el ARNr (que podría enmascarar al nucleoide) mediante hidrólisis ácida o enzimática. Otro buen sistema es emplear el bromuro de etidio, colorante que se intercala en la doble hélice del ADN y que permite visualizar los nucleoides de color anaranjado con un microscopio provisto de luz ultravioleta.
A microscopía electrónica de transmisión
Para evitar artefactos de laboratorio, frecuentes en la fase de fijación para la microscopía electrónica, se recurre a un método de congelación rápida seguida de criosubsitución. Las imágenes muestran al nucleoide como una región más o menos delimitada y libre de ribosomas, dentro del citoplasma, no rodeada de membrana, con muchos lóbulos y un aspecto fibroso. Nuevas técnicas de microscopía electrónica combinada con análisis computerizado de imágenes acaban de revelar que el nucleoide de las células en reposo (en fase estacionaria) se empaqueta de un modo muy ordenado, formando toroides espirales recubiertos de subunidades de una proteína especial.
Métodos de obtención
Para obtener nucleoides “intactos” se recurre a la lisis suave de las células (con detergentes no iónicos) en altas concentraciones de sales (p ej., NaCl) con posterior ultracentrifugación en gradientes de densidad de sacarosa. Si el procedimiento se realiza a 25ºC el nucleoide se aisla relativamente libre de restos de membrana, pero si se hace en frío (4ºC) el nucleoide queda asociado a grandes restos de envueltas.
Estudios moleculares
La biología molecular, especialmente a través de las técnicas de ADN recombinante, permite la caracterización física detallada de los genomas. Muchos genomas de procariotas han sido cartografiados mediante mapas de restricción, pero en la actualidad ya contamos con más de 400 genomas totalmente secuenciados, lo que está permitiendo avances espectaculares en todos los ámbitos de la genética y bioquímica de estos microorganismos.
Los cromosomas aislados muestran una composición de un 60% de ADN, 30% de ARN y 10% de proteínas.
El ADN sigue el modelo clásico de Watson y Crick: dos hebras antiparalelas en doble hélice de 2 nm de diámetro, paso de rosca de 3,4 nm y 10 pares de nucleótidos por cada vuelta de la espiral. La mayor parte de este ADN está en conformación B, aunque existen zonas donde se puede dar la configuración Z.
En la mayor parte de las bacterias este ADN constituye un solo cromosoma circular, cerrado covalentemente (ADN c.c.c.). Existen algunas excepciones, en el sentido de que podemos encontrar cromosomas lineares o incluso más de un grupo de ligamiento (más de un cromosoma):
|
en el género Borrelia el cromosoma es lineal con los extremos cerrados covalentemente (es decir, los extremos forman una especie de bucle de horquilla); | |||||||||
|
bacterias del género Streptomyces también poseen un cromosoma lineal, pero sus extremos contienen secuencias cortas repetidas y están acomplejados con proteínas, lo que recuerda de algún modo los telómeros eucariotas; | |||||||||
|
algunas bacterias parecen poseer dos o más cromosomas:
|
Las bacterias son organismos haploides: poseen un solo cromosoma. Sin embargo, cuando las células bacterianas se encuentran en crecimiento activo, y debido al desfase de la división celular respecto de la replicación, cada individuo puede albergar varias copias de ese cromosoma. Por ejemplo, E. coli puede llegar a 10 copias. Azotobacter puede llegar hasta las 100 copias al final de la fase de crecimiento exponencial. Un caso extremo lo constituye la bacteria gigante Epulopiscium, que aumenta el número de copias en cuatro órdenes de magnitud (¡unas 10.000 veces!), aunque se desconoce el significado de este descomunal “exceso”.
Tamaño del cromosoma
Una bacteria típica, como Escherichia coli posee un cromosoma con 4.700 pares de kilobases (kb). Pero los rangos de tamaño oscilan entre las 700 kb de Mycoplasma genitalium (una bacteria carente de pared y parásita) y las más de 12 000 kb de ciertas bacterias capaces de diferenciación celular y fenómenos de multicelularidad (cianobacterias, actinomicetos).
Organización del cromosoma y de la cromatina bacterianos
Si desplegáramos el cromosoma de E. coli de modo lineal, mediría 1 mm, es decir, 1000 veces más de lo que mide la longitud de la bacteria. Si además tenemos en consideración que el cromosoma ocupa sólo 10% del volumen celular, nos daremos cuenta que debe de estar densamente empaquetado. ¿Cómo es la organización a gran escala de este cromosoma dentro de la célula? Esta es una cuestión que aún desconocemos en todos sus detalles. A continuación exponemos un resumen de lo que se sabe (o sospecha) actualmente: La doble hélice está superenrollada negativamente sobre sí misma. Parte de este superenrollamiento se debe al equilibrio entre la actuación de dos enzimas:
|
ADN girasa, que es un tipo especial de topoisomerasa-II que tiende a introducir superhelicidad negativa; | |
|
ADN topoisomerasa-I, que tiende a relajar la superhelicidad negativa, cortando cada vez en una de las dos cadenas de la doble hélice, pasando la cadena intacta por el hueco, y volviendo a sellar el enlace fosfodiéster que antes rompió. |
Aparte de este superenrollamiento producido por “topoenzimas”, el material genético bacteriano presenta interacciones con una serie de proteínas estructurales. El conjunto de ADN y proteínas estructurales se denomina “cromatina”, pero, nunca aparecen histonas. Salvo en algunas arqueas, esta cromatina procariótica tiene menos densidad de proteínas que la cromatina de eucariotas. Veamos algunas de esas proteínas.
|
En Escherichia coli, existe la proteína básica HU, un heterodímero (HU-a, HU-ß), que presenta cierto parecido con las histonas auténticas (pero sin guardar homología con ellas). No forma auténticos nucleosomas con el ADN. En otras bacterias, existen proteínas homólogas con la HU. En todos los casos se unen débilmente al ADN “normal”, pero en cambio lo hacen con gran afinidad hacia ADN curvado o que forme bucles, induciendo mayores curvaturas en ese ADN. La unión de HU no depende del reconocimiento de ninguna secuencia particular. Parece que su papel no sólo es estructural, sino que también colaboran con otras proteínas en procesos de recombinación homóloga, recombinación específica, reparación del ADN y expresión genética. | |
|
La IHF (llamada así por las iniciales inglesas de factor de hospedador para la integración) es una proteína que reconoce un tipo de secuencia de 13 pares de bases, y que al unirse al surco menor de la doble hélice provoca grandes curvaturas locales en ella. De esta forma colabora en procesos de recombinación específica (lo veremos en la sección de Genética) y de expresión de ciertos genes. | |
|
Recientemente se ha descubierto un importante tipo de proteína estructural para el mantenimiento del cromosoma (SMC). Se trata de un homodímero con forma de "V" en el que cada brazo de la "V" es un monómero. Ambos monómeros interaccionan por la base de la V. Las dos puntas de esa V contienen dominios de unión e hidrólisis del ATP, e interactúan a su vez con una serie de proteínas accesorias relacionadas con la organización y mantenimiento del ADN durante la replicación y segregación de los cromosomas hijos. |
Como se ve, tanto el superenrollamiento como la compactación generada por las proteínas estructurales tienen una notable influencia sobre las funciones del ADN, ya que modulan procesos tan importantes como la replicación, transcripción, recombinación y expresión génica, aunque estamos lejos de conocer exactamente los mecanismos implicados.
El papel biológico del ARN que se detecta en los nucleoides aislados in vitro tampoco está claro. Este no es un ARN especial, sino ARN naciente, y parece servir para mantener “sujetos” los diversos dominios superenrollados.
Ya aludimos antes al descubrimiento reciente de que en la fase estacionaria, cuando las bacterias carecen de fuente de energía y no pueden seguir creciendo, el nucleoide experimenta una reorganización radical, en la que el ADN se arrolla de formando toroides espirales separados entre sí por una capa de subunidades de una proteína especial (Dps), y dando lugar a una macroestructura co-cristalina muy regular. Probablemente esto suponga una adaptación para proteger el ADN sin gastar energía.
Finalmente, otro tema largamente debatido es el modo de asociación del cromosoma con la membrana citoplásmica (¿quizá a través de mesosoma?). Hay indicios de que el cromosoma, y concretamente su origen de replicación (oriC), se encuentra “anclado” a determinadas proteínas de la membrana (proteínas que probablemente estén implicadas en el inicio de la replicación cromosómica). En los cromosomas circulares, la replicación avanza bidireccionalmente desde el sitio oriC, hasta que las dos horquillas se encuentran a 180º de ese lugar, terminándose la replicación.
Aunque en general es adecuado decir que el genoma de los procariotas consta de un solo cromosoma, muchas bacterias poseen, además, uno o varios elementos genéticos accesorios extracromosómicos, a los que denominamos plásmidos.
Se definen como elementos genéticos extracromosómicos con capacidad de replicación autónoma (es decir, constituyen replicones propios). Todos los plásmidos bacterianos estudiados son de ADN de cadena doble. La inmensa mayoría son circulares cerrados covalentemente (c.c.c.) y superenrollados (aunque en Borrelia y algunos Actinomicetos existen plásmidos lineares). Algunos plásmidos poseen, además, la capacidad de integrarse reversiblemente en el cromosoma bacteriano: en esta situación se replican junto con el cromosoma (bajo el control de éste), y reciben el nombre de episomas.
En cuanto al tamaño, existe una amplia gama: desde plásmidos muy pequeños (de unas 2 kb) hasta plásmidos muy grandes (ciertos plásmidos de Pseudomonas tienen 500 kb). Incluso existen “megaplásmidos” en ciertos Rhizobium que llegan a tener 1600 kb. (De hecho, ciertos “megaplásmidos” se ha visto que son imprescindibles, por lo que se han recalificado como cromosomas).
Cada tipo de plásmido tiene un número medio de copias por célula característico. Por regla general, los grandes plásmidos tienen una o dos copias por célula (plásmidos con control estricto de la replicación), mientras que los pequeños suelen estar presentes como varias copias (>10), denominándose plásmidos de control relajado.
En función de que los plásmidos sean o no transmisibles de una bacteria a otra por medio de contactos intercelulares, se pueden distinguir:
|
plásmidos conjugativos (autotransmisibles), que son aquellos que se transfieren entre cepas por medio de fenómenos de conjugación. Algunos de estos plásmidos no sólo se transfieren entre cepas de la misma especie, sino que son capaces de hacerlo entre especies y géneros muy diversos, recibiendo el muy apropiado nombre de plásmidos promiscuos o de amplio espectro de hospedadores, permitiendo transferencia horizontal de información genética entre grupos bacterianos filogenéticamente alejados. | |
|
plásmidos no conjugativos, carentes de esta propiedad de conjugación. Dentro de esta categoría existe un subgrupo, el de los plásmidos movilizables: son aquellos no autotransmisibles que pueden ser transferidos por la acción de un plásmido conjugativo coexistente en la misma bacteria. |
(Ampliaremos el estudio de los plásmidos conjugativos en la sección de Genética bacteriana).
Los plásmidos no suelen ser indispensables para la viabilidad de la bacteria, y muchos de ellos se pierden en ausencia de una presión selectiva. Una bacteria puede ser “curada” de su(s) plásmido(s), es decir, se le pueden eliminar, bien de forma espontánea, bien por una serie de tratamientos en el laboratorio (incubando las bacterias a temperaturas cercanas a la máxima o por agentes químicos como el naranja de acridina, que se insertan entre las bases del ADN). La razón de esta curación de los plásmidos es que esos tratamientos interfieren con su replicación sin afectar a la replicación del cromosoma. Esto hace que en sucesivas divisiones de una bacteria, el plásmido se vaya “diluyendo” en la población resultante.
Los plásmidos no suelen determinar productos esenciales para el crecimiento (por eso son dispensables), pero en la naturaleza parece que resultan favorecidas las bacterias con algún plásmido, quizá porque acarrean ventajas selectivas en determinados ambientes o en determinadas condiciones. Existe una variedad de fenotipos y funciones determinados por plásmidos:
|
Resistencia a antibióticos (plásmidos R; los estudiaremos en la sección de Genética). | |
|
Resistencia a metales pesados (por ejemplo, resistencia a mercurio). | |
|
Plásmidos de virulencia: producción de toxinas, factores de penetración en tejidos, adherencia a tejidos del hospedador, etc., en ciertas bacterias patógenas. | |
|
Producción de bacteriocinas (proteínas tóxicas producidas por bacterias que matan a otras de la misma especie). | |
|
Producción de sideróforos (quelatos para secuestrar iones Fe3+). | |
|
Utilización de determinados azúcares. | |
|
Utilización de hidrocarburos, incluyendo algunos cíclicos recalcitrantes (degradación de tolueno, xileno, alcanfor, etc.) en Pseudomonas. | |
|
Inducción de tumores en plantas (plásmido Ti de Agrobacterium tumefaciens). | |
|
Interacciones simbióticas y fijación de nitrógeno en ciertos Rhizobium. | |
|
etc... |
Obsérvese que la mayor parte de estos fenotipos no están directamente relacionados con el proceso de crecimiento bacteriano, sino que se pueden calificar como funciones para ocupar nichos ecológicos y resistir circunstancias adversas. Los dos fenotipos de “utilización de...(fuente alternativa de C)” nos recuerdan que muchas bacterias están sometidas a períodos alternos de hambrunas y de “festines de comida”: ciertas bacterias, como Pseudomonas se adaptan a los períodos de hambre de las fuentes habituales de C recurriendo a fuentes “exóticas”, como hidrocarburos cíclicos. La información para los enzimas degradativos correspondientes la llevan en plásmidos.
¿Por qué, entonces, si determinadas propiedades conferidas por plásmidos son tan útiles, no han pasado a lo largo de la evolución a ser codificadas por el cromosoma? No podemos contestar a ciencia cierta a esa pregunta, pero cabe especular que resulta ventajoso que algunas poblaciones bacterianas dispongan de ciertos genes en replicones distintos del cromosoma, de modo que los cromosomas no lleguen a ser demasiado grandes. Cuando no hay presión selectiva para los rasgos conferidos por un plásmido, éste se puede perder de parte de la población. Pero cuando existe dicha presión selectiva (p. ej., un antibiótico), sobreviven y se multiplican preferencialmente las bacterias portadoras del plásmido, de modo que en poco tiempo, la población contiene mayoritariamente dicho elemento. Además, como muchos plásmidos son autotransmisibles o movilizables, pueden transferirse a cepas que originalmente carecían de ellos. En resumen, se logra una gran flexibilidad adaptativa sin “cargar” demasiado el cromosoma o replicón principal.
Hay plásmidos que, aunque se pueden evidenciar por métodos físicos (p. ej., electroforesis), no se ha podido demostrar que determinen ningún rasgo fenotípico: tales plásmidos se califican como crípticos, constituyendo un ejemplo de “ADN egoísta” (sólo se mantienen por su capacidad de replicación, sin necesidad de aportar ventaja selectiva clara a la bacteria que los alberga).
Como dijimos, los plásmidos son replicones, es decir, unidades de replicación autónoma, pero ello no significa que codifiquen toda la maquinaria para su propia replicación. De hecho, la mayoría de los plásmidos sólo codifican unas pocas proteínas para este fin, y aprovechan la maquinaria de replicación de la bacteria huésped (ADN polimerasas, ligasas, primasas, etc), que actúa sobre unas secuencias del plásmido denominadas secuencias oriV, donde tiene lugar el inicio de esa replicación. Cada tipo de plásmido se replica por uno de dos principales tipos de mecanismos:
1. Modelo de replicación en q (theta): comienza por la separación de las dos cadenas en el sitio oriV, creando una estructura que se parece a la letra griega q (theta). En algunos plásmidos, la replicación es unidireccional (sólo hay una horquilla de replicación , que avanza a lo largo de la molécula, hasta que vuelve al sitio oriV, en el que las dos moléculas hijas se separan). Otros plásmidos se replican en q por un modelo bidireccional (dos horquillas de replicación de sentidos opuestos, que se encuentran cerca de la mitad de la molécula). La replicación en q es frecuente en plásmidos de bacterias Gram-negativas.
2. Modelo de replicación en s (sigma), o modelo del círculo rodante: una de las dos cadenas es rota a nivel de oriV, de modo que el extremo 3’ suministra el cebador para la replicación de la “cadena adelantada”. La cadena desplazada (cadena “menos”) funciona como cadena retrasada (“lagging”) y debe ser replicada a partir de sitios de cebado especiales. Este tipo de replicación es frecuente en plásmidos de bacterias Gram-positivas.
La regulación del número medio de copias de cada plásmido en cada bacteria es una propiedad característica dependiente de cómo se controla el inicio de replicación, siendo diferentes los mecanismos en los plásmidos de alto número de copias (con control relajado) y en los plásmidos de bajo número de copias (de control estricto).
|
En general, los plásmidos de control relajado tienen mecanismos que inhiben el inicio de replicación sólo cuando el número de copias ha llegado a un cierto nivel. | |
|
En cambio, los plásmidos de control estricto tienen mecanismos que logran que sólo se repliquen una vez o un pequeño número de veces en cada ciclo celular. |
Muchos plásmidos poseen sistemas de reparto (= partición), que tienden a asegurar que una vez que el plásmido ha sido replicado, cada una de las células hijas va a recibir al menos una copia. A falta de un tal sistema, y si el reparto fuera aleatorio, de vez en cuando, las propias fluctuaciones de la segregación aleatoria harían que parte de la progenie no recibiera una copia, con lo que quedaría curada de ese plásmido.
El reparto de las copias a las células hijas se suele deber a las llamadas funciones par, que están cerca de los genes rep y de la zona ori. Se trata de cortas secuencias de ADN que de alguna manera aún no aclarada, debe unirse a alguna zona de la membrana de la bacteria que se duplica durante la división celular, de modo que cada copia, unida a una de esas zonas, se segrega a una célula hija.
Cada plásmido, dentro de la amplia diversidad de los que se conocen, se suele clasificar según el grupo de incompatibilidad al que pertenece (los grupos de incompatibilidad se suelen denominar con las siglas Inc seguidas de una letra mayúscula (por ejemplo IncP, IncFII, etc.). Se dice que dos plásmidos son incompatibles cuando no pueden coexistir establemente en la misma bacteria en ausencia de una presión selectiva permanente. Grupo de incompatibilidad es el conjunto de plásmidos incompatibles entre sí. La incompatibilidad depende del hecho de que los distintos miembros de un mismo grupo poseen el mismo tipo de sistema de control del número de copias y de reparto de dichas copias a las células hijas. En cada célula con dos plásmidos distintos del mismo grupo de incompatibilidad, el número total de copias nA+nB = N (determinado por el sistema de control) no varía, pero por fluctuaciones aleatorias en el reparto, algunas células heredan más copias de uno de los plásmidos que del otro, y tras varias generaciones aparecen células carentes de uno o del otro plásmido (nA = 0 y nB = N, o viceversa).
En los años 70 del pasado siglo se descubrieron en bacterias los primeros ejemplos de ADN “saltarín”, elementos que presentaban la por entonces novedosa propiedad de moverse de un lado a otro de los genomas, pasando desde plásmidos a cromosomas (o viceversa) o desde un plásmido a otro de la misma bacteria (para entendernos, los llamaremos elementos genéticos transponibles “clásicos”, por el simple hecho de que son los que fueron estudiados en primer lugar). Pero desde hace unos 15 años se están descubriendo nuevos tipos de elementos genéticos móviles, que no encajan en el modelo de los clásicos. En conjunto, esta gama de elementos nos están mostrando que en los genomas procarióticos existe una especie de “pool” de ADN con propiedades especiales de movilización, que confieren a veces notables rasgos fenotípicos a las cepas que los poseen,. Además bien por ellos mismos o bien por el hecho de que a veces son transportados entre cepas de la misma especie o de diferentes especies (mediante plásmidos conjugativos y fagos), se pueden transferir de unas bacterias a otras, constituyendo un “pool” compartido de material genético. Y finalmente, estos elementos aportan una notable plasticidad a los genomas, suministrando una base potente sobre la que actúa la selección y por lo tanto la evolución de los procariotas.
Son entidades genéticas discretas que forman parte de cromosomas y de plásmidos, capaces de moverse de un lugar a otro del genoma (pasando a otros lugares del replicón original, o a otros replicones presentes en la misma bacteria). Como estudiaremos oportunamente en la sección de Genética, esta capacidad de transponerse se debe a un tipo de recombinación específica denominado transposición. Existen varias clases de mecanismos de transposición, pero muchas de ellas suponen la simultánea replicación del elemento, de modo que queda una copia en el sitio original y aparece una copia nueva en la nueva localización.
Los elementos genéticos móviles acarrean una serie de reorganizaciones en los genomas: inversiones de segmentos genéticos, delecciones, cointegraciones entre dos replicones, etc. Hay que resaltar que estos elementos no son replicones, es decir, no poseen la capacidad de replicación autónoma que hemos visto en cromosoma y plásmidos, sino que son partes integrantes de los genomas de muchas bacterias, confiriendo plasticidad genética sobre la que pueden actuar mecanismos evolutivos. Esto hace que los genomas bacterianos, y sobre todo los plásmidos, sean relativamente dinámicos y maleables a escala evolutiva.
![]()
Como dijimos, en los últimos años se han descrito nuevos tipos de elementos móviles que no se ajustan a los clásicos. Algunos de ellos poseen funciones simultáneamente de plásmidos y transposones, otros tienen mezclas de rasgos de fagos y de plásmidos, etc. Describiremos brevemente algunos de ellos:
|
Uno de los primeros tipos de “nuevos” elementos son los llamados transposones conjugativos, que presentan la propiedad de promover su diseminación de una bacteria a otra por conjugación. | |||||||||||||||||
|
Los transposones movilizables consisten en un transposón que posee un sitio oriT similar al de ciertos plásmidos conjugativos, y que puede ser transferido “pasivamente” a otra célula mediante un plásmido conjugativo co-residente (es decir, presente en la misma célula de origen). | |||||||||||||||||
|
Las islas genómicas son elementos discretos de ADN con tamaños de entre 10 y 500 kb, presentes en ciertas cepas de una especie pero ausentes en otras, y que confieren notables ventajas adaptativas (para ocupar nichos ecológicos). Muchas islas genómicas se asemejan a enormes transposones atípicos, que se insertan preferentemente en o cerca de ciertos genes cromosómicos (como p. ej., genes de ARNt), que están enmarcados por secuencias cortas repetidas (similares a las que usan ciertos fagos, como el fago λ), y que de hecho codifican una enzima de tipo integrasa como la del propio fago λ. Algunas de estas islas a su vez pueden transferirse activamente a otras células debido a que poseen también genes parecidos a los de los plásmidos conjugativos. A su vez, las islas genómicas son de varios tipos en función de la ventaja adaptativa que confieren a las cepas que las albergan:
|
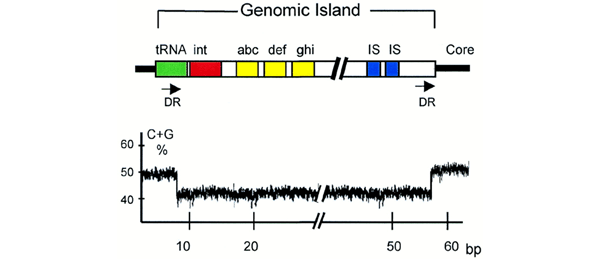
En resumidas cuentas: los elementos móviles permiten que los genomas procarióticos sean relativamente “modulares” y “maleables”, y habida cuenta de que por sí mismos o por el efecto de plásmidos y fagos se pueden movilizar entre cepas y especies diferentes, suministran mecanismos rápidos de cambio evolutivo. En el caso de la adquisición de islas genómicas, observa que el fenotipo de la bacteria cambia espectacularmente (p. ej., de no patógena a patógena, de no simbiótica a simbiótica, etc), por lo que este fenómeno se ha calificado como de “evolución por saltos cuánticos”.
_______________________
Antes de terminar esta parte del tema sobre el ADN procariótico, debemos decir que la mayor parte del genoma bacteriano consta de genes y conjuntos de genes (y a diferencia de eucariotas, existe poco ADN “no génico” y pocas secuencias cortas repetidas sin función clara). La mayoría de los genes codifican proteínas, bien sean estructurales o enzimáticas. La “ruta” de expresión por la que la secuencia de un gen se decodifica hasta proteína consta de dos fases: transcripción y traducción.
Un concepto muy importante a tener ya en cuenta, y que distingue la organización genética procariótica de la eucariótica es que los genes bacterianos relacionados con una misma función genética suelen estar agrupados de forma contigua, formando una unidad de transcripción y de regulación genética que se denomina operón, bajo el control unitario de unas secuencias de ADN colocadas en el lado 5’, entre las que siempre existe una llamada promotor, que es el lugar de entrada de la ARN polimerasa.
La ausencia de membrana nuclear en los procariotas hace que la traducción de un mensaje ocurra mientras ese mensaje aún no ha terminado de formarse: tan pronto como ha comenzado la transcripción de un operón, los ribosomas se unen al extremo 5' del mensajero naciente, y da comienzo enseguida la traducción del mensaje. Esto constituye otro rasgo distintivo de la expresión genética en procariotas: la transcripción y la traducción están estrechamente acopladas. Igualmente característico de las eubacterias es el hecho de que el primer aminoácido que se incorpora no es la metionina, sino la N-formil-metionina.
Como ya vimos, en las micrografías electrónicas de cortes finos de bacterias el citoplasma aparece muy granulado, correspondiendo estos granos a los 10.000 a 15.000 ribosomas que posee cada célula (dependiendo de la fase de crecimiento).
El ribosoma es probablemente el complejo supramacromolecular más estudiado, a pesar de lo cual, y debido a su extraordinaria complejidad, aún reserva una buena cantidad de aspectos no totalmente comprendidos.
Nos referiremos a la composición y estructura del ribosoma eubacteriano (concretamente, de la especie en que está mejor estudiado: Escherichia coli, por supuesto).
El ribosoma está compuesto de un 63% de ARN (que a su vez representa más del 90% del ARN total de la bacteria) y un 37% de proteínas. El ribosoma eubacteriano posee un coeficiente de sedimentación de 70S, frente al de 80S de los ribosomas citoplásmicos eucarióticos. Bajando la concentración de iones Mg++ cada ribosoma se disocia en sus dos subunidades: la pequeña (30S) y la grande (50S). In vivo esta disociación ocurre cada vez que se completa la síntesis de una molécula de proteína, para volver a unirse las dos subunidades al inicio del mensaje de otro gen.
Múltiples técnicas moleculares (algunas relativamente recientes) permiten desentrañar aspectos de los ribosomas:
|
difracción de rayos X; | |
|
difracción de neutrones; | |
|
inmunoelectromicroscopia. |
Subunidad pequeña (30S)
Papeles:
|
Está implicada principalmente en decodificar la información del ARNm. | |
|
Contiene los sitios de unión para los ARNt cargados. | |
|
Tiene un papel central en el inicio de la traducción. |
Composición y estructura:
|
Contiene un solo tipo de ARN: el ARNr 16S, con una característica estructura secundaria con zonas de emparejamiento intracatenario (de cadena doble) y bucles. | |
|
Posee 21 tipos de proteínas, denominadas S1, S2 ... S21. Las posiciones relativas de algunas de estas proteínas han podido ser “cartografiadas” en el conjunto de la estructura de la subunidad 30S por las técnicas citadas arriba. |
Subunidad grande (50S)
Papeles:
|
Interviene principalmente en la formación del enlace peptídico entre el aminoácido situado en el sitio A (ligado a su ARNt) y el péptido naciente (unido a un ARNt) del sitio P. |
Composición y estructura:
|
Posee dos tipos de ARN: ARNr 23S y ARNr 5S, cada uno con su correspondiente y peculiar estructura secundaria (En general, los ARNr presentan abundantes zonas de emparejamientos intracatenarios y bucles de cadena sencilla). | |
|
Contiene 32 tipos de proteínas diferentes, denominadas L1 ... L32. La L7 y la L12 tienen la misma secuencia, pero la L7 está modificada químicamente en su extremo amino por unión con un radical acetilo. Con excepción de L7/L12, que están presentes en 4 copias cada una, las demás aportan una sola molécula cada una a la subunidad grande. Véase en la figura la localización de algunas de estas moléculas dentro de la estructura global. |
La secuencia primaria y la estructura secundaria de los ARNr están muy conservados evolutivamente: hay pocas diferencias entre bacterias muy alejadas desde el punto de vista filogenético (¿Podría el alumno explicar el porqué de esta notable conservación química y estructural?). Parece ser que el papel principal de los ARNr es suministrar un “núcleo” sobre el que se van ensamblando ordenadamente las proteínas ribosómicas, interaccionando con sitios específicos de los ARN y con otras proteínas.
Recientemente se están acumulando pruebas de que el ARNr pueda jugar, además, algún papel funcional, aparte del estructural:
|
El ARNr 16S, además de unirse por su extremo 3' con la secuencia de Shine-Dalgarno del extremo 5' del ARNm, parece representar un papel en la terminación de la síntesis de proteínas. | |
|
El ARNr 23S tiene un papel en la elongación, interaccionando con factores EF. Incluso existe la sospecha de que es una ribozima necesaria para la actividad peptidil-transferasa. |
Aún no tenemos un cuadro completo de cómo coordina el ribosoma la compleja serie de reacciones que forman parte de la traducción: unión y liberación de los ARNt, transpeptidación, movimientos coordinados del molde de ARNm y del péptido naciente, etc. Sin embargo, el intenso estudio al que se están sometiendo el ribosoma y el proceso de la traducción desde los años 60 ha permitido notables profundizaciones. El alumno de biológicas puede recurrir a los conocimientos que se imparten en otras asignaturas de la Licenciatura (Bioquímica, Genética, Biología Molecular) y a alguno de los excelentes textos modernos.
El sitio de entrada del ribosoma al ARNm es la secuencia de Shine-Dalgarno (S-D), cerca del extremo 5’ del mensajero, de unas 9 pb, y complementaria de extremo 3’ del ARNr 16S (de la subunidad pequeña del ribosoma). Esta secuencia de S-D está situada dentro de una región más amplia de secuencia no aleatoria que abarca desde la base –20 a la +13. También es importante la distancia entre la S-D y el codón iniciador.
|
La formación del complejo iniciador con la subunidad ·30 S requiere los factores de iniciación IF1, IF2 e IF3, el ARNm y un ARNt especial, cargado normalmente con el aminoácido N-formil-metionina. (El grupo formilo bloquea el grupo amino de ese aminoácido iniciador, de modo que obliga a que la síntesis proceda sólo en una dirección). |
No siempre el codón de inicio es AUG (met), sino que a veces lo son GUG (Val) y UUG (Phe).
|
Entonces se añade la subunidad 50S, formándose el ribosoma 70S, y liberándose los factores de iniciacion | |
|
El ARNt fMet se encuentra en el sitio P del ribosoma. Al lado de él, en el sitio A, ha entrado el ARNt correspondiente al siguiente codón, cargado con su aminonácido. En la fase de traslocación, el ribosoma, junto con el factor de elongación EF-G forma un enlace peptídico entre estos dos aminoácidos, forzando la liberación del f-Met de su ARNt y moviéndose el ribosoma un codón adelante. Las dos moléculas de ARNt permanecen unidas al ribosoma, pero ahora el ARNt descargado se sitúa en el sito E (exit=salida), y el ARNt con la cadena naciente (por ahora de 2 aminoácidos) queda en el sitio P. | |
|
Un nuevo ciclo comienza: entra el tecer ARNt cargado al sitio A, y en presencia del factor de elongación EF-Tu, libera al ARNt descargado del sitio E. | |
|
Y continúan los ciclos de elongación (por traslocación) hasta llegar al codón de parada. Hay tres codones que no tienen sentido (no equivalen a ningún aminoácido): UAA; UAG, UGA. Cuando uno de estos codones llega al sitio A, no hay ARNt que lo traduzca, y la cadena proteica queda liberada del ribosoma. |
Como los procariotas suelen tener mensajes policistrónicos, cuando un ribosoma llega al final de un gen, se puede mover hasta encontrar otro codón iniciador, pero si el espacio intercistrónico es muy grande, se puede disociar el ribosoma, de modo que las subunidades deben reensamblarse para continuar la lectura del mensaje.
Hasta hace poco se pensaba que el polipéptido naciente adquiría espontáneamente su configuración funcional al ser sintetizado en el ribosoma. Pero hoy se sabe que tanto el correcto plegamiento de las proteínas como su adecuado ensamblaje en complejos requiere el concurso de unas proteínas especiales, conocidas como proteínas celadoras o “carabinas moleculares”, debido a que su papel es vigilar y eventualmente corregir el plegamiento. (Se está imponiendo, para denominarlas, la castellanización de su nombre inglés: chaperonas, procedente de chaperones).
Estas proteínas están presentes en todos los seres vivos. Algunas de ellas se inducen ante determinados estrés ambientales; de hecho se descubrieron como proteínas inducibles ante un aumento de temperatura, por lo que se denominaron como proteínas del choque térmico ( con su acrónimo inglés Hsp).
En Escherichia coli se han estudiado especialmente la DnaK, DnaJ, GrpE, GroEL y GroES. Parece que actúan de un modo secuencial:
1. Cuando el ribosoma aún está sintetizando la proteína y ya ha “asomado” la primera porción del polipéptido, se une la DnaJ a esa parte del polipéptido aún sin plegar.
2. Enseguida se une la DnaK, que ha formado antes un complejo con el ATP, y se produce hidrólisis de este ATP (pero quedando el ADP unido a DnaK), lo que aumenta la afinidad de DnaK por el polipéptido sin plegar.
3. Cuando se ha sintetizado toda la proteína, la GrpE se une al complejo polipéptido-DnaJ-DnaK-ADP, liberando el ADP.
4. Nuevas moléculas de ATP se unen ahora a DnaK, con lo que se facilita la disociación de las proteínas DnaK y DnaJ respecto del polipéptido. En este momento, muchos polipéptidos pueden haber adquirido su configuración final, pero otros necesitan aún un paso final de “refinamiento”:
5. El polipéptido pasa al canal interior formado por GroEL y GroES, que catalizan (con gasto de ATP) la correcta isomerización, de modo que la proteína adquiere su plegamiento nativo biológicamente activo.
Son acúmulos de sustancias orgánicas o inorgánicas, rodeadas o no de una envuelta limitante de naturaleza proteínica, que se originan dentro del citoplasma bajo determinadas condiciones de crecimiento. Constituyen reservas de fuentes de C o N (inclusiones orgánicas) y de P o S (inclusiones inorgánicas).
Estudiaremos:
1) Inclusiones orgánicas:
a) inclusiones polisacarídicas
b) gránulos de poli-ß-hidroxibutírico (o, en general de poli-ß-hidroxialcanoatos)
c) inclusiones de hidrocarburos
d) gránulos de cianoficina
2) Inclusiones inorgánicas:
a) gránulos de polifosfato
b) glóbulos de azufre
Son acumulaciones de a(1à4) glucanos, con ramificaciones en a(1à6), principalmente almidón o glucógeno (según especies), que se depositan de modo más o menos uniforme por todo el citoplasma cuando determinadas bacterias crecen en medios con limitación de fuente de N, pero donde aún sean abundantes las fuentes de C y energía. En esta situación, se detiene prácticamente la síntesis de proteínas y de ácidos nucleicos, y la mayor parte del C asimilado se convierte rápidamente en estos materiales de reserva. Cuando a estas células las pasamos a un medio rico en N, pero carente de fuente de C, estas inclusiones se usan como fuente interna de C para la síntesis de ácidos nucleicos y proteínas.
Estas inclusiones actúan, pues, como sistemas de almacenamiento de carbono osmóticamente inertes (la célula puede albergar grandes cantidades de glucosa que, si estuvieran como moléculas libres dentro del citoplasma, podrían tener efectos osmóticos muy negativos).
Para observarlas se recurre a la tinción con una solución de I2 + IK (como el lugol)
|
glucógeno: aparece de color pardo-rojizo; | |
|
almidón (amilopectina): color azul |
Los gránulos de poli-b-hidroxibutírico son acúmulos del poliéster del ácido ß-hidroxibutírico (= 3-hidroxibutírico), rodeados de una envuelta proteínica, y que al igual que en el caso anterior, se producen en ciertas bacterias como reserva osmóticamente inerte de C en condiciones de hambre de N. Además de la protección osmótica, estos gránulos suponen la ventaja de neutralizar un metabolito ácido (el grupo carboxilo de cada unidad de ß-hidroxibutírico desaparece como tal, al intervenir en el enlace éster con la siguiente unidad). En las especies de Bacillus constituye la fuente de carbono y energía al inicio de la esporulación. Una función semejante parece implicada a la hora del enquistamiento de Azotobacter.
Una célula puede contener de 8 a 12 de estos gránulos, que miden unos 0.2-0.7 mm de diámetro, y que van provistos de una envuelta proteica de unos 3-4 nm de grosor. Pueden llegar a representar el 80% en peso de la célula.
A diferencia de los acúmulos de polisacáridos, los gránulos de PHB son visibles a microscopio óptico en fresco, debido a su elevado índice de refringencia. Se tiñen bien mediante Negro-Sudán.
En los últimos años está quedando patente que los gránulos descritos de PHB son un ejemplo de una clase más amplia de gránulos de poli-ß-hidroxi-alcanoatos.
Por ejemplo: cuando determinadas especies de Pseudomonas crecen en n-octano como fuente de carbono, se acumula un polímero de ésteres del ácido ß-hidroxi-octanoico, con una función metabólica semejante a la del PHB.
Ciertas cepas de Ralstonia eutropha, cuando crecen en glucosa y propiónico producen copolímeros aleatorios de unidades de b-hidroxibutírico y b-hidroxivalérico (=3-hidroxipentanoico).
Existen interesantes perspectivas de aprovechamiento económico de estos polímeros, ya que los PHA se comportan como excelentes termoplásticos biodegradables. Por ejemplo, la empresa británica ICI tiene patentados procesos industriales para fabricar PHA donde casi el 90% de las unidades son de hidroxivalérico, que da un polímero flexible comercializado con el nombre de Biopol ®. Los polímeros a base de 4- o 5-hidroxibutírico y 3-hidroxibutírico son más largos, más elásticos y más biodegradables (se han empleado en la fabricación de envases)
Son acúmulos de reserva (con envuelta proteinica) de los hidrocarburos que determinadas bacterias (especialmente Actinomicetos y relacionados) usan como fuente de C.
Muchas cianobacterias (Oxifotobacterias) acumulan grandes gránulos refringentes de reservas nitrogenadas cuando se acercan a la fase estacionaria de crecimiento. Estos gránulos de cianoficina son acúmulos de un copolímero de arginina y aspártico: consta de un núcleo de poliaspártico, en el que todos los carboxilos de las cadenas laterales están unidos con L-arginina. Su síntesis no está basada en el mecanismo habitual en ribosomas, ya que no se ve inhibida por el cloramfenicol.
Se denominan también gránulos de volutina o metacromáticos. El nombre de “metacromáticos” alude al efecto metacromático (cambio de color): cuando se tiñen con los colorantes básicos azul de toluidina o azul de metileno envejecido, se colorean de rojo. A microscopio electrónico aparecen muy densos a los electrones.
Son acúmulos de polifosfato, polímeros lineales del ortofosfato, de longitud variable (por término medio, unas 500 unidades), que representan un modo osmóticamente inerte de almacenar fosfato. Parece ser que la parte central de estos gránulos constituye un núcleo formado por lípidos y proteínas. En algunos casos pueden constituir una fuente de energía, en sustitución del ATP (¿se trata en este caso de una especie de “fósil bioquímico?”).
Se acumulan cuando algún otro nutriente distinto del fosfato se hace escaso (sobre todo cuando va desapareciendo el sulfato). En estas condiciones se detiene la síntesis de los ácidos nucleicos, y la volutina se acumula a la espera de su utilización para esta síntesis de nucleicos, cuando aparezca el nutriente originalmente limitante.
Las inclusiones de S aparecen en dos grupos de bacterias que usan sulfuro de hidrógeno (SH2):
|
las bacterias purpúreas del azufre (que usan el SH2 como donador de electrones para la fotosíntesis); | |
|
bacterias filamentosas no fotosintéticas como Beggiatoa, Thiomargarita o Thiothrix, que lo usan como donador de electrones para sus oxidaciones. |
En ambos casos, el sulfuro de hidrógeno es oxidado a azufre elemental (S0), que en el citoplasma se acumula como glóbulos muy refringentes y rodeados de envuelta proteínica. Estos glóbulos son transitorios, ya que el S0 se reutiliza por oxidación hasta sulfato, cuando en el medio se agota el sulfuro.
Acúmulos grandes, densos y refringentes de sales insolubles de calcio (sobre todo carbonatos) que aparecen en algunas bacterias (como Achromatium), cuyo papel parece consistir en mantenerlas en el fondo de los lagos y ríos.
Son estructuras supramacromoleculares, en forma de cilindros o bastones, adosadas a la superficie de la membrana tilacoidal de las Cianobacterias, confiriendo a ésta un típico aspecto “granuloso” en las micrografías electrónicas.
Como se puede ver en el esquema, están constituidas por pilas de discos a base de ficobiliproteínas, cromoproteínas que sirven como “antenas” para la captación de luz en la fotosíntesis de estos procariotas. Los grupos cromóforos son: ficocianinas, aloficocianinas y ficoeritrina. Como veremos oportunamente, la disposición ordenada de los distintos pigmentos tiene un papel central en la “canalización” de la energía de la luz hacia los centros de reacción (ubicados ya en plena membrana tilacoidal) donde se localizan los complejos fotosintéticos proteínas-clorofilas.
Como ya dijimos, en procariotas no existen por regla general orgánulos citoplásmicos rodeados por unidad de membrana. Las únicas excepciones están constituidas por los tilacoides de las Oxifotobacterias, ya estudiados en el capítulo anterior. En algunos grupos bacterianos se pueden encontrar orgánulos citoplásmicos no rodeados por unidad de membrana (o sea, sin bicapa lipídica). Muchos de ellos presentan envueltas basadas en subunidades de proteínas:
|
carboxisomas | |
|
vacuolas de gas | |
|
clorosomas | |
|
magnetosomas. |
Estructuras presentes en bacterias fotoautotrofas (Cianobacterias y ciertas bacterias purpúreas) y quimioautotrofas (nitrificantes, Thiobacillus), de apariencia poliédrica con tendencia a esférica. Su diámetro oscila entre 50 y 500 nm, y están rodeadas de envuelta monocapa proteinica de unos 3,5 nm. El interior tiene aspecto granular, debido a la acumulación de la enzima ribulosa-bifosfato-carboxilasa (RuBisCo, la carboxidismutasa, el enzima clave en el ciclo de Calvin de asimilación de CO2). Aunque se pensó que eran los sitos de fijación del CO2, parece más bien que se trata de reservas de dicha enzima.
Son orgánulos muy refringentes al microscopio óptico, que al electrónico muestran una estructura a base de agrupaciones regulares de vesículas de gas. Cada vesícula tiene una forma de cilindro bicónico (200-1000 nm de longitud y unos 70 nm de diámetro), rodeado de una monocapa de unidades globulares de proteína ensambladas helicoidalmente que dan un aspecto a bandas (“costillas”). Esta envuelta es impermeable al agua, pero permeable a los gases, por lo que la composición y concentración del gas dentro de la vesícula depende de las que existan en el medio. Conforme se sintetizan y ensamblan las vesículas, el agua va siendo eliminada del interior.
Las vesículas de gas están constituidas por dos tipos de proteína: la mayoritaria GvpA es una pequeña proteína rígida y muy hidrófoba. Su rigidez está en la base del hecho de que las vesículas y vacuolas de gas aguanten presiones externas. La proteína minoritaria GvpC tiene como función reforzar las vesículas de gas.
La función de estas vacuolas es mantener un grado de flotabilidad óptimo en los hábitats acuáticos a las bacterias que las poseen, permitiéndoles alcanzar la profundidad adecuada para su modo de vida (según los casos, para obtener una intensidad adecuada de luz, concentración óptima de oxígeno o de otros nutrientes).
Las vacuolas de gas son muy frecuentes en eubacterias fototrofas (Cianobacterias y bacterias fotosintétiocas purpúreas y verdes); también se dan en algunas arqueas (Halobacterium, algunas metanógenas) y en bacterias prostecadas (Ancalomicrobium, Prosthecomicrobium).
Son vesículas oblongas situadas por debajo de la membrana citoplásmica, que contienen los pigmentos antena de las bacterias fotosintéticas verdes (antigua familia Chlorobiaceae). Son invisibles a microscopía óptica; miden 100-150 nm de longitud y unos 50 nm de anchura, estando rodeadas de una monocapa de proteínas. Se disponen por debajo de la membrana citoplásmica, sin estar en continuidad con ella, aunque en muchos casos aparecen conectadas a través de un pedúnculo de naturaleza no lipídica.
Son orgánulos sensores del campo magnético terrestre, que aparecen en ciertas bacterias acuáticas flageladas microaerófilas o anaerobias (p. ej., en Aquaspirillum magnetotacticum). Consisten en cristales homogéneos de magnetita (Fe3O4), de formas cubo-octaédricas o de prisma hexagonal delimitados por una envuelta proteínica. Los diversos cristales suelen disponerse en filas paralelas al eje longitudinal de la bacteria, o en otras agrupaciones regulares de varios unidades, hasta varias decenas.
Fueron descubiertas en 1975, y se sabe que permiten la orientación magnética a las bacterias que las poseen (bacterias magnetotácticas), determinando la orientación de su natación. En el hemisferio Norte, el campo magnético está orientado hacia abajo, y en el sur hacia arriba. Las bacterias magnetotácticas del hemisferio septentrional se orientan al N, y las del meridional, al S. Por consiguiente, cuando las bacterias son removidas de los fondos donde viven, por magnetotaxia pueden volver al fondo, que es donde encuentran las concentraciones de oxígeno adecuadas para su modo de vida.
![]()
![]()



