
Actualizado el miércoles, 15 febrero 2006
1 LA ENDOSPORA BACTERIANA Y LA ESPORULACIÓN
1.1 INTRODUCCIÓN A LA ESPORULACIÓN BACTERIANA
1.2 OBSERVACIÓN DE LAS ESPORAS
1.3 ESTRUCTURA Y COMPOSICION QUIMICA DE LA ENDOSPORA
1.4 DESENCADENAMIENTO DE LA ESPORULACIÓN
1.6 GENÉTICA MOLECULAR DE LA ESPORULACIÓN
1.8 PROPIEDADES BIOLÓGICAS DE LAS ESPORAS: SU FUNDAMENTO
1.9 GERMINACIÓN DE LA ENDOSPORA
3 DIFERENCIACIONES EN LOS ACTINOMICETOS
5 DIFERENCIACIONES EN CIANOBACTERIAS
![]()
Hasta ahora hemos abordado la estructura y funciones de las diversas partes de la célula procariota. Para terminar esta sección del programa, en este capítula vamos a tratar de algunos casos de diferenciación celular en bacterias, es decir, procesos por los que a partir de células “normales” (vegetativas) se producen formas celulares especializadas. Algunas de ellas son formas de reposo, capaces de aguantar mucho tiempo en ausencia de nutrientes (caso de la endospora). Muchas de ellas pueden resistir circunstancias ambientales adversas (la endospora es, de hecho, la forma bacteriana más resistente a calor, desecación, luz UV, etc).
Algunas especies de bacterias Gram positivas (principalmente de los géneros Bacillus, Clostridium, Sporosarcina y Thermoactinomyces), disponen de una notable estrategia adaptativa cuando se ven sometidas a privación de nutrientes en su medio ambiente: si los niveles de fuentes de C, N, o P caen por debajo de un umbral, esto constituye una señal a la célula de que se avecina un largo período de privación de nutrientes. Entonces, la célula se implica en una serie de complejos cambios genéticos, metabólicos, estructurales, etc. (proceso de esporulación o esporogénesis), que conducen a la diferenciación, en el interior de la célula vegetativa original, de una célula durmiente (la endospora). La célula-madre (o sea, la célula vegetativa original que generó la endospora) finalmente se autolisa, liberando la espora, que es capaz de permanecer en estado criptobiótico, durmiente, varios decenios, -incluso siglos. Las esporas son fácilmente diseminadas por el aire; cuando caen en medios ricos en nutrientes, se desencadena su germinación, se reinicia la actividad metabólica, de modo que cada espora genera una nueva célula vegetativa, capaz de divisón binaria, etc.
O sea, la esporulación se puede considerar como un proceso de supervivencia “en última instancia”, la “última carta” que se juegan ciertas bacterias Gram positivas cuando se enfrentan a condiciones severas de hambre de nutrientes.
Significado adaptativo: Estas bacterias suelen vivir en medios nutricionalmente pobres (suelos, hierba seca, etc.). La esporulación es un proceso muy refinado que ha aparecido evolutivamente en una línea filogenética de bacterias Gram-positivas, y que les permite sobrevivir largos períodos de carencia nutricional. (Atención: NO son estrictamente formas de resistencia).
Las endosporas son formas de reposo (y no formas reproductivas), que representan una etapa del ciclo de vida de ciertas bacterias, y que se caracterizan por una estructura peculiar, diferenciada respecto de las células vegetativas, por un estado metabólico prácticamente detenido, y por una elevada resistencia a agentes agresivos ambientales.
La esporulación bacteriana es un sistema modelo donde se pueden estudiar, con relativa sencillez, determinados problemas biológicos más generales: ¿qué bases moleculares y genéticas tiene la diferenciación de un tipo de célula a otro tipo distinto?, ¿cómo se regula el proceso en función del tiempo y del espacio?, ¿cómo se “reparten los papeles” los tipos celulares implicados?, etc. Por esta razón, presenta un enorme interés para los biólogos, interesados en escudriñar las bases íntimas de este tipo de importantes cuestiones, tan frecuentes en los organismos superiores.
Al microscopio óptico, en fresco (sin teñir), aparecen como cuerpos esféricos, ovoides e incluso en algunas especies, cilíndricos, muy refringentes, libres, o aún incluidos en la célula vegetativa (célula madre).
Esporangio = célula madre + espora
El tamaño relativo de la espora, y su situación en el esporangio, son criterios taxonómicos importantes en las bacterias esporulantes.
|
Según que el diámetro de la espora sea o no mayor que el de la célula vegetativa:
| |||||||
|
Según la localización de la espora dentro del esporangio:
| |||||||
|
Los esporangios deformantes de Clostridium son característicos:
|
Tinción:
No se tiñen por los colorantes normales. Hay que forzar por calor y/o mordientes (por ejemplo, tras teñir reforzadamente con fuchsina, resisten decoloración por alcohol-ClH). Otra tinción muy empleada es la reforzada con verde de malaquita (que es la que el alumno realiza en nuestras prácticas de laboratorio).
Partes que comprende la endospora:
|
Protoplasto o núcleo (“core”, en inglés), con la membrana citoplásmica de la espora (membrana esporal interna). | |
|
Pared de la espora (= Germen de la pared de la futura célula vegetativa). | |
|
Corteza o córtex, rodeado externamente de la membrana esporal externa. | |
|
Cubiertas | |
|
Exosporio (no universal: las esporas de algunas especies carecen de él). |
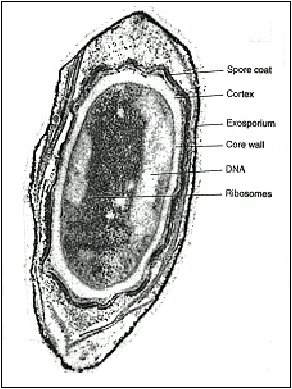 |
Micrografía electrónica que muestra las partes principales de una endospora |
El citoplasma de la espora está muy deshidratado. Sus componentes están inmovilizados en una matriz de quelatos de iones Ca++ y ácido dipicolínico.
El citoplasma de la espora contiene altas concentraciones de ion Ca++ (1-3% del peso seco de la espora), y de ácido dipicolínico (DPA) (10% en peso); ambos están formando un quelato, llamado dipicolinato cálcico (DPC), una sustancia exclusiva de las esporas bacterianas.
Papel del DPC: Como veremos, es una “reserva de Ca2+”, que durante la esporulación secuestra este ión, lo que tendrá un papel importante en la deshidratación del protoplasto; durante la germinación, la liberación del Ca2+ será fundamental en este proceso.
El protoplasto contiene un cromosoma completo, condensado, y todos los componentes indispensables para reiniciar el crecimiento vegetativo cuando la espora germine, pero carece de muchos componentes típicos de la célula vegetativa:
|
posee una pequeña dotación de ribosomas, junto con componentes estables de la maquinaria de síntesis de proteínas (ARNt, cofactores, etc.) | |
|
ARN polimerasa. | |
|
nucleósidos mono- y difosfatados, pero no NTP (no hay ATP) | |
|
carece de componentes inestables: no hay ARNm, ni enzimas biosintéticas, ni aminoácidos ni bases nitrogenadas libres, ni cofactores reducidos (NADH; CoA) | |
|
pero en cambio, existe una gran cantidad de una serie de pequeñas proteínas especiales (llamadas SASP, del inglés “small acid-soluble proteins”). Cuando se produzca la germinación, estas proteínas serán hidrolizadas rápidamente, y los aminoácidos resultantes serán empleados en la síntesis de nuevas proteínas necesarias en la germinación y crecimiento ulterior. Además, las SASPs están acomplejando al ADN (induciendo en él un cambio conformacional hacia la rara forma A, en vez de la B habitual), protegiéndolo del daño por luz UV. | |
|
la “moneda energética” de la espora es el 3-fosfoglicerato, otra molécula estable del protoplasto, que será convertido rápidamente a 2-fosfoglicerato, y de él a PEP (fosfo-enol-pirúvico) al germinar la espora (el PEP es donador de P para generar ATP). |
Rodeando al protoplasto está la membrana citoplásmica (membrana interna de la espora), una bicapa lipídica carente de fluidez, posiblemente como resultado de su estructura policristalina.
Situación: inmediatamente por encima de la membrana interna de la espora (ver micrografía a microscopio electrónico, o el esquema de la ultraestructura).
Composición: a base de un peptidoglucano (PG) similar al de la célula vegetativa, con sus característicos enlaces entre los tetrapéptidos.
Funciones: al germinar la espora, dará lugar a la pared celular de la nueva célula vegetativa, confiriéndole, mientras tanto, resistencia osmótica.
Origen: se sintetiza a partir de la prespora, atravesando los precursores la membrana interior que hemos citado arriba.
(Obsérvese su aspecto a microscopio electrónico: transparente a los electrones, relativamente gruesa, a base de láminas concéntricas).
Composición: un peptidoglucano (PG) especial, diferente en composición al PG de la célula vegetativa:
|
30% del NAM tiene tetrapéptidos normales, pero el grado de entrecruzamiento es muy bajo (6%). | |
|
15% del NAM tiene solo la L-ala inicial, en lugar de tetrapétido. | |
|
55% de una modificación del ácido murámico (lactama del ácido murámico), producida por condensación del -COOH lactilo con el -NH2, para formar la lactama correspondiente). |
Origen: Se sintetiza a partir de la célula madre, con sus intermediarios ensamblados a nivel de la membrana externa que rodea a la corteza.
Propiedades de la corteza:
1) Tiene un bajo grado de puentes entre tetrapéptidos (sólo un 6%). Ello condiciona:
a) una estructura más laxa, floja y flexible que el peptidoglucano normal, capaz de expandirse en ausencia de cationes que neutralicen sus grupos negativos (sobre todo del mDAP y del glutámico de los tetrapéptidos). Esto es la base de su apariencia de gel.
b) su rápida autolisis, durante la germinación de la espora.
2) la lactama del murámico condiciona una gran resistencia a la lisozima.
La corteza está limitada por la membrana esporal externa, procedente de invaginación de la membrana citoplásmica de la célula vegetativa madre. Posee una polaridad opuesta a la de la membrana esporal interna.
Composición y estructura: la composición depende de las especies, pero en general, a base de una o varias proteínas de tipo queratina, todas muy ricas en cisteína y en aminoácidos hidrófobos, y que llegan a constituir el 60% en peso seco de la espora. Buena parte de la cisteína se añade post-traduccionalmente, por modificación de la proteína inmadura. A microscopio electrónico se puede comprobar el gran grosor (volumen) de las cubiertas, y su aspecto relativmante denso a los electrones.
Propiedades: son muy insolubles e impermeables, e impiden la entrada de numerosos agentes químicos, incluyendo sustancias tóxicas. La abundancia de puentes S-S las hace muy compactas y muy estables químicamente.
En B. cereus es una estructura membranosa transparente, a modo de saco cerrado, delgado y flojo, envolviendo al resto de la espora de una forma “suelta”, despegado en principio respecto de las cubiertas (tras la liberación de la espora, al perderse el contenido acuoso que había entre exosporio y cubiertas, aquel se pega a éstas).
En B. subtilis y B. megaterium, el exosporio está envolviendo a la espora de una forma más estrecha, estableciendo algunos contactos físicos con la superfie de las cubiertas.
Composición química: mezcla de proteínas, polisacáridos complejos, y lípidos.
Propiedades: muy resistente a enzimas proteolíticas, lo que sugiere (pero no prueba directamente) que el exosporio puede representar algún papel como barrera de defensa externa de la espora.
Para que se produzca la esporulación, se necesitan dos condiciones previas:
1) los cultivos bacterianos han de estar en buenas condiciones;
2) cuando cesa el crecimiento exponencial, la mayoría de las células entran en esporulación, aunque el proceso no es sincrónico (hay desfases entre unas células y otras), de modo que en un lapso de tiempo relativamente breve (5,5-8 horas en Bacillus subtilis) casi todo el cultivo aparece en forma de esporas, habiendo desaparecido las células vegetativas. La división celular típica de la fase de crecimiento exponencial y la esporulación son procesos mutuamente excluyentes.
¿Qué estímulo es el responsable de desencadenar la esporulación? Lo que detiene el crecimiento de estas bacterias y dispara la esporulación es un estado de inanición, de carencia de nutrientes. El nutriente limitante que puede desencadenar la esporulación puede ser: la fuente de carbono, la fuente de nitrógeno o incluso la carencia de fosfatos.
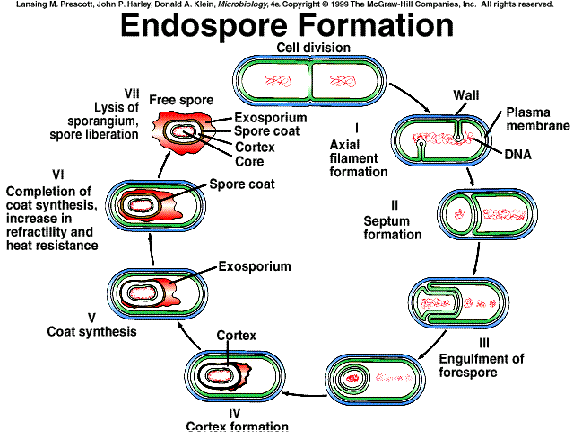
Vamos a estudiar la esporulación en Bacillus subtilis o en B. megaterium, dos de las bacterias esporuladas mejor investigadas, y que completan la esporulación al cabo de unas 7-8 horas de su inicio, a 37ºC. Se pueden distinguir siete fases consecutivas (además de la fase “0” anterior a la esporulación). Cada fase se denota con un número romano, y su duración en horas se expresa como subíndice de t, p. ej., t2. Como veremos enseguida, cada fase se distingue por un(os) evento(s) citológico(s) peculiar(es) y por una serie de cambios bioquímicos propios:
Fase 0 (célula vegetativa):
Al final del periodo de crecimiento exponencial, la célula vegetativa contiene dos cromosomas.
Fase I (t0-t1):
|
El material genético (los dos cromosomas) se condensa constituyendo un filamento axial ancho, que ocupa el centro de la célula, según su eje longitudinal. Cada nucleoide está unido a uno de los extremos de la célula. | |
|
En vez de iniciarse una división celular simétrica (con septo central), se forman dos espículas de la pared celular cerca de los polos hacia el interior, rodeadas de la correspondiente invaginación de la membrana citoplásmica. Cada espícula está señalando una zona donde se está formando el anillo de FtsZ (proteína contráctil de tipo tubulina: véase el tema 5) | |
|
Se sintetizan y se liberan al medio antibióticos y varias exoenzimas. Una serie de proteasas se encargan del turnover de proteínas intracelulares, cuyos aminoácidos constituyentes serán empleados para sintetizar nuevas proteínas específicas de la esporulación. |
Fase II (t1-t2):
|
Se termina por formar un septo transversal acéntrico, cerca de un polo de la célula, por invaginación de la membrana citoplásmica, y deposición de nuevo peptidoglucano entre las dos membranas adyacentes. | |||||
|
Cada nucleoide queda segregado en uno de los dos compartimentos que se han formado:
| |||||
|
En esta fase continúa la síntesis de los antibióticos y de las exoenzimas (serina-proteasas, metalo-proteasas, ribonucleasas, a-amilasa, etc). |
Fase III (t2-t3):
|
Independización del protoplasto de la preespora respecto de la célula madre. Para ello, lo primero que ocurre es la degradación selectiva del peptidoglucano del septo que se había depositado en la fase II (esta degradación comienza por el centro, es decir, por donde se había cerrrado, y sigue hacia la periferia, y en ella intervienen los productos de varios genes). Entonces, la membrana citoplásmica de la célula madre va creciendo unidireccionalmente alrededor de la preespora, hasta que ésta queda libre en el citoplasma del esporangio. | |
|
Obsérvese que el citoplasma de la preespora queda rodeado por dos membranas de polaridad opuesta: la interior tiene la polaridad normal, pero la exterior, derivada del crecimiento de la membrana de la célula madre, tiene polaridad invertida | |
|
A partir de esta fase el turnover (renovación) de proteínas sólo tendrá lugar en la célula madre, deteniéndose en el compartimento de la preespora. |
Fase IV (t3-t4):
|
Se forma casi por completo la corteza de la espora, por deposición de peptidoglucano entre las dos membranas. Sin embargo este PG aún no está “maduro” (no tiene todavía las características que estudiamos en el apartado 1.3.3). | |
|
También se deposita el peptidoglucano de la pared celular (que constituye el germen de la pared celular de la futura célula vegetativa). | |
|
Coincidiendo con esto, la preespora adquiere su aspecto refráctil al microscopio óptico en fresco. | |
|
Comienza la síntesis del ácido dipicolínico (DPA), así como la acumulación de Ca2+. | |
|
En las bacterias que lo poseen, en esta fase comienza la síntesis del exosporio. |
Fase V (t4-t5,5):
|
Los materiales de las cubiertas (que se habían ido sintetizando durante las fases II y III en el compartimento de la célula madre), comienzan a depositarse por fuera de la membrana esporal externa. | |
|
Al final de esta fase se adquiere la resistencia al octanol. | |
|
Continúa la acumulación de DPA, que sigue secuestrando iones Ca2+ (formación del quelato de dipicolinato cálcico, DPC). |
Fase VI (t5,5-t7):
|
La preespora madura hasta espora. | |
|
Madura la corteza (para generar el característico PG cortical, más laxo, con su bajo porcentaje de entrecruzamientos -por actuación de endopeptidasas- y la lactama del NAM). | |
|
Maduran las cubiertas. | |
|
El citoplasma de la espora se hace más homogéneo y más denso a los electrones. | |
|
Por una serie de razones aún no totalmente aclaradas, la espora se hace resistente al calor y al cloroformo. | |
|
Se adquiere resistencia a las radiaciones ultravioleta (UV). | |
|
Se adquiere resistencia a la lisozima. |
Fase VII (t7-t8):
|
Liberación de la espora madura por autolisis de la célula madre. | |
|
las bacterias que han producido exosporio, cuando la espora sale al exterior, este exosporio pierde su contenido de citoplasma (procedente de la célula madre), quedando como un saco vacío y plegado, unido a las cubiertas. | |
|
La parte superior de la figura muestra un gráfico de los principales cambios que acabamos de comentar, expresados en función del tiempo transcurrido desde el inicio de la esporulación. |
Las células pueden detener el proceso de la esporulación si durante las 4 primeras fases se les suministra el nutriente que estaba limitando y que fue responsable del desencadenamiento (efecto de desbloqueo metabólico). Pero a partir de la fase V el aporte de nutrientes no tiene ya ese efecto inhibidor: el proceso de la esporulación ya es irreversible, y se dice que la célula está comprometida (= obligada) a esporular.
El proceso de la esporulación es muy ordenado en el tiempo y en el espacio. En su base genética existe un conjunto de varios operones específicos de la esporulación (operones spo), sometidos a un finísimo control genético espacial y temporal. El conjunto de estos operones (que se encuentran en cuatro zonas distintas del cromosoma), se denomina esporulón, y comprende más de 125 genes. Las principales características de los operones del esporulón son:
|
Los operones están inactivos durante el crecimiento vegetativo; | |
|
se van activando de modo ordenado y secuencial, para luego ser reprimidos (una vez han actuado); | |
|
están sometidos a interacciones pleiotrópicas unidireccionales; | |
|
existe una “comunicación” regulatoria entre el compartimento de la espora y de la célula madre (interacciones espaciales); | |
|
dentro de los mecanismos genéticos implicados en la expresión y regulación de los distintos operones correspondientes a fases distintas de la esporulación, un papel muy importante es el jugado por las subunidades sigma (s) alternativas de la ARN-polimerasa. |
Algunas bacterias esporuladas, como Bacillus thuringiensis, B. popiliae y algunas especies de Clostridium, forman cristales proteicos en el esporangio simultáneamente a la formación de la endospora: son los llamados cuerpos parasporales.
Cada célula madre exhibe una sola inclusión, que se puede presentar libre en el citoplasma, o bien englobada en el exosporio de la espora. Los cuerpos parasporales pueden ser amorfos, pero los más típicos son pseudocristales octaédricos (bipiramidales). Están compuestos de la agregación regular de subunidades de una glucoproteína de unos 120 kDa, sintetizada durante la fase IV.
La función de estos cuerpos parasporales en la esporulación es desconocida (e incluso puede que de hecho, no tenga tal tipo de función). Bajo condiciones alcalinas, se disuelven y la proteína se convierte en una poderosa toxina para larvas de lepidópteros y otros insectos, cuando las ingieren vía oral (pero no por vía parenteral). Veamos cómo se produce el proceso de la toxicidad:
1. La oruga come materia vegetal que lleva bacterias esporuladas. El cuerpo parasporal se libera junto con la espora, cuando la célula madre se autolisa.
2. Los cuerpos parasporales se disuelven en el tracto digestivo de la oruga (que es alcalino), la proteína sufre proteolisis, lo que la transforma en una toxina.
3. Esta toxina altera la permeabilidad del epitelio intestinal de la oruga, de modo que los líquidos alcalinos del intestino pasan a la hemolinfa, que incrementa su pH por encima de 8, lo cual termina provocando la parálisis rápida del insecto, y finalmente su muerte.
El significado biológico más probable de este fenómeno es que la producción de cuerpos parasporales sea una variante de la esporulación que evolucionó durante la adaptación de estas bacterias a sus nichos ecológicos, como una manera de asegurar la germinación de las endosporas: la oruga ingiere los cristales junto con las esporas. Los cristales parasporales matan a la oruga, que se pudre. La oruga muerta y en putrefacción aseguraría un entorno adecuado en nutrientes para que se alimentaran y multiplicaran las células vegetativas que surgieran de la germinación de esas esporas.
Desde hace tiempo ciertos grupos de agricultores vienen usando inóculos de bacterias esporuladas de las especies productoras de cuerpos parasporales para rociar sus plantas y protegerlas de insectos: estamos ante un auténtico insecticida biológico, biodegradable, selectivo hacia las plagas e inofensivo para los seres superiores.
La moderna Ingeniería Genética ha logrado insertar y expresar genes codificadores de las toxinas parasporales de Bacillus thuringiensis en plantas de cultivo. Hoy día existen millones de hectáreas de cultivo de plantas transgénicas "Bt" (sobre todo algodón, maíz, y patata) que dependen menos de los insecticidas químicos merced a estos genes bacterianos (aunque ello no ha evitado las críticas de ciertos grupos ecologistas opuestos por sistema a todo lo que suene a manipulación genética por técnicas de ADN recombinante).
Las endosporas son células en estado de dormancia, con una bajísima tasa metabólica (hipometabolia, la menor que existe en el mundo vivo), y capaces de conservar su vitalidad durante larguísimos períodos. Son muy resistentes a la acción de diversos agentes químicos (octanol, cloroformo) y físicos (altas temperaturas, congelación, desecación, radiaciones).
1) Hipometabolia: Poseen la más baja tasa respiratoria de todos los seres vivos. Por ello son capaces de sobrevivir en ausencia de nutrientes durante largos períodos de tiempo.
2) Dormancia: Esta propiedad se refiere al hecho de que la espora tiene una gran inercia a los sustratos exógenos. Como veremos, la espora sólo perderá la dormancia cuando se haya activado para la germinación.
3) Resistencia al calor: Las esporas de ciertas especies resisten el calor húmedo de 120oC durante 10 min, lo cual condiciona los parámetros para esterilizar materiales (véase el capítulo 13 titulado “Acción de agentes físicos sobre las bacterias”). Esta elevada resistencia a las altas temperaturas es un subproducto de los cambios evolutivos que condujeron a la deshidratación como medio para lograr la hipometabolia y la dormancia. Por lo tanto, la deshidratación es la clave de las anteriores propiedades. (En cambio, como dijimos antes, al menos parte de la resistencia al calor seco, es decir, en ausencia de vapor de agua, se debe a las proteínas SASP, que protegen al ADN de los daños oxidativos de este tipo de calor).
4) Deshidratación: El muy bajo contenido en agua de la espora (0.3 g de agua/g de peso seco frente a los 3-4 g de agua/g de peso seco de la célula vegetativa) hace que la espora sea muy refráctil al microscopio óptico en fresco. Ello condiciona las propiedades 1, 2 y 3. ¿Cuál es el mecanismo de la deshidratación, base a su vez de tantas propiedades biológicas de las endosporas? El tema es aún objeto de debate. Vamos a exponer brevemente una hipótesis (1989), según la cual habría 3 etapas en la consecución de la espora deshidratada y resistente al calor:
a) En las fases III y IV el ácido dipicolínico va entrando al protoplasto de la prespora. Simultáneamente el Ca2+ entra desde el exterior a la célula madre por transporte activo, y de ella al protoplasto de la prespora por difusión facilitada. Se forma el correspondiente quelato de dipicolinato cálcico (DPC). Este es un proceso con retroalimentación positiva, ya que conforme el Ca2+ queda “secuestrado” en forma de quelato (lo que significa que no hay de hecho iones Ca2+ libres en el interior de la prespora), se facilita más la entrada de mayores cantidades de Ca2+. Parte del Ca2+ y de otros cationes se acomplejan también con macromoléculas del protoplasto. Todo ello redunda en que la corteza se queda sin cationes; por lo tanto, no hay posibilidad de neutralizar las cargas negativas del peptidoglucano cortical à las cargas negativas de la corteza se repelen à se produce la expansión del peptidoglucano cortical à en su expansión, la corteza se “topa” con las cubiertas rígidas, por lo que presiona sobre el protoplasto à sale más agua del protoplasto.
b) Resultado final: termoestabilización (= termorresistencia) del protoplasto. La gran pérdida de agua hace que los diversos compuestos del protoplasto se compacten entre sí, lo que facilita sus interacciones con los cationes. El resultado es la formación de un gel a base de una matriz constituida por complejos supramoleculares, macromoléculas y pequeñas moléculas densamente empaquetados, unidos entre sí e inmovilizados. Parece ser que esta es la base principal de la enorme resistencia al calor húmedo por parte de la endospora.
5) Resistencia a los rayos UV: Parece que depende de varios componentes:
a) absorción de luz UV por las cubiertas;
b) por el DPC;
c) pero cada vez está más claro que las proteínas SASP tienen un papel central en esta resistencia a los UV. Como ya dijimos, las SAPS de tipo α/β acomplejan al ADN y favorecen su configuración de tipo A, lo cual a su vez provoca un cambio en su fotoquímica (ver apartado siguiente);
d) cambio en la fotoquímica del ADN de la espora: se puede explicar parcialmente por la misma deshidratación del protoplasto, que impide que se formen dímeros de pirimidina entre pirimidinas adyacentes de una misma cadena del ADN. Como veremos en el capítulo 13, estos dímeros de pirimidina constituyen el principal tipo de fotoproductos generados por rayos UV en el ADN de la célula vegetativa, base de la mayor parte de los efectos deletéreos de estas radiaciones. Por otro lado, las SASP de tipo α/β que acomplejan el ADN hacen que en la espora la luz UV provoque otro tipo de alteración, llamada fotoproducto de la espora (que consiste en 5-timinil-5,6-dihidrotimina) que se produce en menor cantidad. Aunque la acumulación de fotoproductos de la espora puede ser igual de letal que la de los dímeros de pirimidina, cuando germina la espora se pone inmediatamente en marcha un eficiente sistema de reparación específico de ese fotoproducto.
6) resistencia a agentes químicos: La resistencia de la endospora a agentes como octanol, cloroformo, etc. se debe a la impermeabilidad de las cubiertas, gracias a su gran grosor y su peculiar composición a base de proteínas ricas en aminoácidos hidrófobos y con abundantes puentes disulfuro (cistinas). La resistencia a la lisozima se debe por un lado a la propia impermeabilidad de las cubiertas, y a la resistencia de la corteza.
La germinación es el proceso por el cual una espora se convierte al estado vegetativo. Es mucho más rápida que la esporulación (dura unos 90 minutos). Podemos considerar en ella cuatro etapas, según el modelo de Foster y Johnstone (1990):
1. preactivación
2. activación
3. iniciación (o germinación en sentido estricto)
4. crecimiento ulterior (entrada en fase vegetativa)
Antes de que la espora esté en condiciones de germinar se requiere que sus cubiertas se alteren. En la naturaleza esto ocurre por erosión por envejecimiento progresivo. Artificialmente, en laboratorio, se puede recurrir a algún procedimiento para alterar esas cubiertas:
|
tratando las esporas a altas temperaturas, pero inferiores a su inactivación (100oC durante unos minutos); | |
|
por radiaciones ionizantes; | |
|
por pH bajos; | |
|
por tratamiento con sustancias que posean grupos -SH libres (p. ej., mercaptoetanol). |
La activación es una etapa aún reversible, desencadenada por un agente químico externo (germinante) presente en el medio. Este agente es variable según las especies:
|
iones inorgánicos (Mn2+, Mg2+); | |
|
L-alanina en B. subtilis; | |
|
glucosa u otros azúcares; | |
|
adenina u otras bases nitrogenadas. |
El germinante es detectado por un receptor alostérico a nivel de la membrana esporal interna. Una vez que dicho receptor se activa, adquiere una capacidad proteolítica específica que le permite romper una proenzima que hasta ese momento se encontraba unida covalentemente al peptidoglucano de la corteza. La enzima resultante reconoce la lactama del NAM y comienza a hidrolizar el peptidoglucano cortical. La consecuencia es que comienza a entrar agua al protoplasto de la espora, por lo que la espora pierde su característica refringencia, y se comienza a perder la resistencia al calor.
Durante toda esta etapa el metabolismo está aún latente.
En esta etapa la germinación se hace ya irreversible, y se rompe definitivamente el estado de dormancia, si bien el metabolismo es endógeno (no depende todavía de sustancias externas). Los principales acontecimientos bioquímicos son:
|
se pierde DPA, lo que supone pérdida de Ca++; | |
|
este Ca++ pasa al córtex, donde neutraliza las cargas negativas à se favorece la rehidratación del protoplasto y su hinchamiento, favorecido por la concomitante contracción del córtex, mientras continúa y se completa rápidamente la hidrólisis del PG cortical; | |
|
el 3-fosfoglicérico (3-PG) se convierte en 2-PG, y éste en PEP, que a su vez dona su fosfato de alta energía para producir ATP; | |
|
las pequeñas proteínas SASPs se hidrolizan por una proteasa específica que hasta ese momento estaba inactiva. De este modo los aminoácidos constituyentes de las SASPs se reutilizan para la síntesis de nuevas proteínas por parte de la pequeña dotación de ribosomas y demás moléculas accesorias; | |
|
La ARN polimerasa comienza a sintetizar ARN (comienza la transcripción de genes vegetativos). |
Aparece ya el metabolismo exógeno, de modo que la espora puede tomar nutrientes del exterior y metabolizarlos. Los eventos bioquímicos y estructurales más notorios son:
|
se sintetiza ADN; | |
|
el protoplasto crece aún más; | |
|
la pared de la espora sirve como cebador (germen) para la producción de la pared de la célula vegetativa naciente; | |
|
la célula vegetativa sale por rotura de las cubiertas, que puede ser de tipo polar o ecuatorial. Hay que aclarar que al salir, esta célula vegetativa se tiñe como Gram-negativa, y sólo adquirirá su típica grampositividad después de la primera división. |
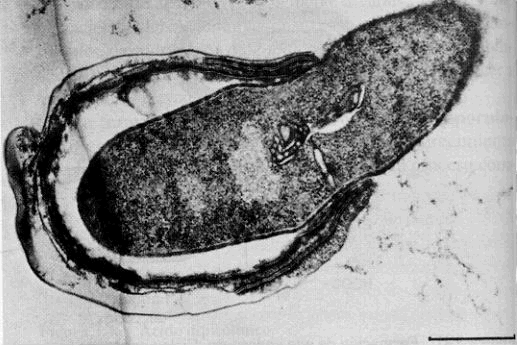 |
Salida de la nueva célula vegetativa al final de la germinación. Observa la rotura de las cubiertas |
Determinadas bacterias (Methylosinum, Rhodomicrobium) forman esporas reproductivas por gemaciones sucesivas al final de sus prostecas. Estas exosporas poseen una envuelta a base de pared rodeada de una cápsula o cubierta gruesa.
Los actinomicetos constituyen un grupo amplio y complejo de bacterias Gram positivas con tendencia a un tipo de crecimiento micelial y un estilo de vida similar a los hongos. Muchos de los taxones de Actinomicetos y bacterias relacionadas poseen células diferenciadas de tipo reproductivo, genéricamente conocidas como esporas. Estudiaremos brevemente la diferenciación en dos géneros típicos.
Género Actinoplanes
Las especies de este género producen micelios vegetativos de sustrato (subterráneos). Algunos de estos micelios generan hifas verticales que sobresalen a la superficie. El extremo de cada una de estas hifas se diferencia para constituir un saco llamado esporangio, que se fragmenta en un conjunto de esporas móviles (flageladas) llamadas zigosporas o esporangiosporas.
Género Streptomyces
Forma un micelio de sustrato ramificado, interrumpido de vez en cuando por pared transversal (son por lo tanto organismos cenocíticos, es decir, el segmento de micelio limitado por dos tabiques sucesivos contienen varios cuerpos nucleares). Cuando hay limitación de nutrientes se comienza a formar un micelio aéreo a partir de ramificaciones de las hifas subterráneas. En los extremos de algunas de estas hifas aéreas las células se diferencian en cadenas de esporas. Durante la formación de estos micelios aéreos y de las esporas la población de micelios subterráneos sufre una lisis masiva.
Propiedades de las esporas de los estreptomicetos:
|
la pared celular de la espora es más gruesa que la de la célula vegetativa; | |
|
no hay cambio cualitativo en el peptidoglucano; | |
|
no hay córtex ni cubiertas; | |
|
son muy hidrofóbicas: se resisten a ser suspendidas en agua. Esto parece que se debe a una vaina que rodea a la pared celular, compuesta a base de túbulos auto-ensamblables. | |
|
resisten más al calor y a la desecación en comparación a las células vegetativas, pero menos que las endosporas. | |
|
son metabólicamente durmientes (células en reposo). |
Son células que se producen en algunas especies por engrosamiento de la pared celular de la célula vegetativa, por deposición de nuevos materiales externamente a la membrana citoplásmica, al mismo tiempo que se acumulan materiales de reserva en el citoplasma. Poseen metabolismo endógeno, y resisten al calor, a la desecación y a agentes químicos más que la correspondiente célula vegetativa (pero menos que las endosporas).
Ejemplos:
|
Quistes de Azotobacter y Bdellovibrio. | |
|
Microquistes de Mixobacterias, llamados mixosporas: sus envueltas constan de una corteza, rodeada de cubiertas (interna y externa). Estas cubiertas se componen de una glucoproteína muy rica en polisacáridos. |
En las cianobacterias filamentosas (que forman tricomas) se pueden observar dos tipos principales de células diferenciadas a partir de las vegetativas: heteroquistes y acinetos.
Heteroquistes (= heterocistes)
Son células de término, sin función reproductiva, especializadas en la fijación de nitrógeno molecular (N2), de mayor tamaño que las células vegetativas.
La conexión entre células vegetativas y heteroquistes se establece a través de un estrangulamiento de los polos de éstas. La unión está atravesada por una serie de finos canales llamados microplasmodesmos, que reducen al mínimo el intercambio de sustancias entre ambas células.
Por fuera de la pared celular (que es de tipo Gram-negativo), existen tres cubiertas:
|
una capa laminada interna a base de glucolípidos exclusivos de cianobacterias; | |
|
una capa homogénea central a base de polisacáridos; | |
|
una capa fibrosa externa, también polisacarídica, pero menos compactada. |
Estas tres capas evitan la difusión del O2 al interior del heteroquiste, lo que representa uno de los mecanismos para la protección de la nitrogenasa (complejo enzimático que reduce el N2 a NH4+, y que es muy sensible al oxígeno).
Pero el heteroquiste dispone de más “estrategias” para la protección de la nitrogenasa:
|
aparte de los microplasmodesmos, en las uniones con las células vegetativas adyacentes se forman sendos “tapones” de cianoficina; | |
|
los tilacoides se disponen como un retículo polar y periférico, y carecen de ficobilisomas, por lo que no pueden realizar la fase II de la fotosíntesis. Por lo tanto, los heteroquistes no generan oxígeno, ya que sólo realizan la fotofosforilación cíclica. |
Acinetos (=aquinetos)
Son formas de reposo que se originan a partir de células vegetativas, por acumulación de nuevas capas de materiales polisacarídicos por fuera de la pared celular, y por formación de acúmulos de reserva en el citoplasma.
Resisten más que las células vegetativas los períodos de desecación y de congelación, pero no al calor.
Cuando las condiciones ambientales mejoran, se producen sucesivas divisiones transversales en el acineto, que finalmente se convierte en un filamento más corto y menos grueso que los tricomas, llamado hormogonio.



