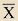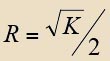|
PATRONES Y TENDENCIAS EN LA ENCEFALIZACIÓN DEL GÉNERO HOMO PATTERNS AND TRENDS IN HOMO GENUS’ ENCEPHALIZATION Alexia SERRANO RAMOS* Resumen Palabras Claves Abstract Key words
En la actualidad, un cerebro humano adulto pesa entre 1000 y 2000 gramos, con un promedio de 1330 gramos. El cerebro supone un 2% del peso total del cuerpo aunque consume aproximadamente un 20% de la energía metabólica basal en reposo. Lamentablemente el cerebro no fosiliza. Las evidencias que nos permiten aproximarnos al estudio de la evolución del cerebro provienen de la paleoneurología, que estudia los moldes endocraneales y aporta información sobre el volumen endocraneal así como algunos detalles de la morfología del cerebro que quedan impresas en el endocráneo. La Neuroanatomía comparada se dedica al estudio de seres vivos y de ahí se extrapola información para las especies extintas a partir de analogías. La Arqueología nos permite recuperar tanto los restos paleoantropológicos como las evidencias de la conducta de los homínidos. La investigación sobre el proceso de encefalización del género Homo posee una larga tradición de estudio, desde los mismos inicios de la paleoantropología. En parte porque el cráneo es uno de los elementos que mejor queda preservado en el registro fósil y porque por largo tiempo, el incremento del cerebro fue considerado el motor de la evolución humana. Este aumento del volumen endocraneal es una de las características que se han utilizado, con mayor o menor grado de aceptación, para distinguir al género Homo del resto de homínidos (vg. australopithecinos), junto a la reducción del aparato masticatorio, la morfología de la mano y la capacidad de fabricar útiles de piedra tallada. Sin embargo, el incremento del cerebro no explica por sí solo lo original de nuestras capacidades cognitivas. De esta manera, el proceso de encefalización debe entenderse como un mosaico de eventos evolutivos en el que se interrelacionan el incremento del tamaño cerebral así como diferentes reorganizaciones de los componentes cerebrales. Con todo, a día de hoy no hay pruebas de que, a nivel intraespecífico, un cerebro mayor signifique mayor inteligencia ni se ha demostrado ninguna relación convincente entre el tamaño del cerebro y el comportamiento, a excepción de los casos patológicos (microcefalia e hidrocefalia). Lo que sí es cierto es que un mayor volumen del cerebro implica mayor número de neuronas e interconectores. Si atendemos al peso absoluto del cerebro, las ballenas (7,800 g para el cachalote – Physeler macrocephalus) y los elefantes (7,500 g para el elefante africano – Loxodonta africana) tendrían que considerarse los animales más encefalizados. Sin embargo, hay que tener presente que el cerebro es el encargado de controlar los procesos vegetativos a la par que su mantenimiento supone un alto coste metabólico. A principios del siglo XX se describió la “ley de la disarmonía” (THOMPSON 1917) que describe el fenómeno por el cual el tamaño del cerebro aumenta por debajo de lo que lo hace el tamaño del cuerpo. Así, en los años 70 del siglo XX Jerison (1973) acuñó el término de encefalización tratando de cuantificar el número extra de neuronas no implicadas en los procesos vegetativos del cuerpo. El incremento cerebral también se ha puesto en relación con el tamaño del cuerpo y su variablidad, por ejemplo aplicando a su estudio la regla ecogeográfica de Bergmann (1847) que predice un aumento de la masa corporal para aquellos individuos de una especie ampliamente distribuida que habitan en latitudes más alejadas del Ecuador como una posible adaptación para disminuir el área de superficie frente al tamaño corporal en los climas más fríos. Con la explosión en las neurociencias a finales del siglo XX el cerebro se empezó a considerar un todo integrado, y se pasó de la consideración de una única variable neural a una perspectiva multivariada de presiones selectivas operando en el tamaño del cerebro, incluyendo niveles de comportamiento. Milton (1981, 1983) relacionaba dicho aumento con la búsqueda de alimentos. Por otro lado, algunos investigadores se centraron en la disponibilidad de energía que permita mantener un cerebro grande. Aiello & Wheeler (1995) basaban en un cambio de dieta con mayor ingesta de proteínas animales que habría permitido reducir el gasto energético del sistema digestivo. Matarese & La Cava (2004) explican dicha disponibilidad a partir de que se rebajase el gasto del sistema inmunológico de aquellos homínidos que emigrasen fuera de las regiones ecuatoriales, en cuyas zonas se da una menor cantidad de parásitos dañinos (LOW 1990). En otra dirección, surgieron hipótesis que relacionaban el aumento del cerebro con la presión social. Alexander (1989) conjugaba la reducción de la mortalidad y una mayor eficiencia en la obtención de recursos naturales que desembocaría en un aumento de población y llevando de esta manera, la presión social a la cabeza del cambio evolutivo del cerebro. Holloway (1967, 1975) también esbozó lo que se denominó posteriormente la “hipótesis del cerebro social” (DURBAN 1998) donde se remarca la importancia del procesado de información de tipo social como un importante factor en el tamaño del cerebro, en concreto del neurocórtex. En la misma línea, Bailey & Geary (2009) conjugaron el incremento de densidad de población con el paleoclima. Un último grupo de hipótesis sobre el aumento del cerebro se pueden agrupar en torno a modelos ecológicos/ nichos ecológicos/ y la variabilidad paleoclimática. A partir de la relación de mayores volúmenes endocraneales y de coeficientes de encefalización en animales con complejos sistemas de recolección/caza (BARTON 1996) se plantearon hipótesis enfocadas a las ventajas de la caza y otras adaptaciones, como la fabricación y uso de herramientas, que facilitarían la obtención de recuros (KAPLAN et al. 2000). Otros remarcan la relación entre la variabilidad climática ligada a medioambientes impredecibles y la ventaja adaptativa de poseer un cerebro mayor (ASH & GALLUP 2007), a partir de la hipótesis de selección – variablidad (POTTS 2001) por la que mayoritariamente, la selección natural se da más por fluctuaciones medioambientales que en un entorno de constantes medioambientales. Generar nuevos conocimientos en el ámbito de la evolución humana puede ser conseguido a través de dos estrategias. Por un lado se pueden obtener más datos. Esto puede lograrse con el hallazgo de nuevas evidencias fósiles, pero también a través de nuevas técnicas que permitan obtener más información de los viejos fósiles. Por otro lado podemos mejorar la forma en que los datos de los que disponemos son analizados (WOOD 2005). Las ideas (ligadas a una metodología) nos permiten avanzar, tanto o más, que los nuevos fósiles.
De esta forma, el presente estudio tiene como objetivos: (1) evaluar estadísticamente el cambio en el volumen endocraneal, la masa corporal y el coeficiente de encefalización en los diferentes taxones del género Homo; (2) estudiar el grado de solapamiento existente para cada variable entre los distintos taxones; (3) testar tanto la influencia del tiempo, (4) como de los principios ecogeográficos (regla de Bergmann) con las principales del proceso de encefalización para tratar de explicar la variabilidad del tamaño del cerebro/cuero que hallamos en el registro fósil.
Toda la información relativa a los individuos fósiles fue recogida de la literatura científica, mayormente de Ruff et al. (1997), De Miguel & Hennerberg (2001) y Ash & Gallup (2007). Todas las observaciones fueron introducidas en una base de datos para, posteriormente, analizarlas estadísticamente. Se ha utilizado el paquete estadístico SPSS y la hoja de cálculo de Excel. Se seleccionaron aquellos individuos adultos cuyos restos paleoantropológicos permitiesen conocer o bien el volumen endocraneal o la estimación de la masa corporal, es decir, aquellos individuos fósiles de los que se han conservado el cráneo o una parte de éste que permitiese su reconstrucción, o bien el fémur o la pelvis para estimar la masa corporal siguiendo a Ruff et al. (1994). La muestra total es de 323 individuos, de los cuales 294 pertenecen al Pleistoceno y 29 al Holoceno. Las variables consideradas en este estudio son: (1) TAXONOMÍA, siguiendo una propuesta conservadora o lumper en la que se tienen en cuenta los caracteres discretos como los adecuados para la clasificación taxonómica (JIMÉNEZ ARENAS et al. 2011). Se han considerado 5 grupos principales: (a) los habilinos (comprende a Homo habilis, H. rudolfensis y la población de Dmanisi), (b) los erectinos del Pleistoceno inferior (Homo ergaster y Homo erectus), (c) los erectinos del Pleistoceno medio (Homo erectus, Homo heidelbergensis y Homo mauritanicus), (d) los neandertales (Homo neanderthalensis), (e) los humanos anatómicamente modernos del Pleistoceno (Homo sapiens) y por último se consideró un grupo adicional (f) de humanos anatómicamente modernos del Holoceno (integrado en este caso por individuos del pueblo Pecos, Nuevo México). (2) DATACIÓN, las fechas manejadas están expresadas en miles de años (kyr). Cuando diversas dataciones se ofrecían para un mismo individuo fósil se ha optado por elegir aquellas absolutas frente a relativas, o aquella fecha absoluta que con mayor frecuencia aparece en la literatura. No se han tenido en cuenta las recientes dataciones con carbono 14 por infiltración, que ahora mismo tienden a envejecer a los fósiles puesto que aún no ha sido aplicado a un porcentaje significativo. (3) VOLUMEN ENDOCRANEAL (VE en adelante), expresado en centímetros cúbicos (cc). Supone la medición de la capacidad craneal, que contiene tanto el cerebro como los tejidos y vasos sanguíneos que lo envuelven, y como cualquier variable métrica está sujeta a errores de medición, paliados en mayor o menor medida, según el método utilizado para su medición (métodos “de relleno”, estimaciones a partir de reconstrucciones físicas y ahora más comúnmente virtuales, estimaciones a partir de fórmulas derivadas de medidas externas del cráneo – LEE & PEARSON 1901-). Cuando se disponía de diversas mediciones de VE para un mismo individuo se consideró el promedio de éstas, a no ser que el método de medición haya sufrido alguna mejora sustancial (primando una reconstrucción virtual de una parte del endocráneo frente a la estimación resultante de una variable lineal). (4) ESTIMACIÓN DE LA MASA CORPORAL (MC en adelante), expresada en kilogramos (kg), resulta una variable más complicada ya que no existe ninguna medida esquelética ue refleje directamente el peso del cuerpo de un individuo. Para poder realizar estimaciones fiables de la masa corporal es necesario emplear huesos o partes de éstos que jueguen un papel importante en la función de soportar el peso corporal. Se han escogido los datos que presentan Ruff et al. (1997) a partir de la utilización de dos métodos independientes que permiten estimar la masa corporal y que presentan resultados concordantes al aplicarlos a los individuos pleistocenos. No se han tenido en cuenta las estimaciones de MC a partir de la altura orbital propuesta por Aiello & Wood (1994) porque los errores en las estimaciones son superiores al 25%. (5) MASA CEREBRAL (MCe) expresada en gramos (g), es propiamente el peso del cerebro, que suele ser entre un 8 y un 12% menor que el volumen endocraneal (HOLLOWAY 2009). Esta variable se ha calculado siguiendo la fórmula presentada en Ruff et al. (1997): Masa Cerebral = (volumen endocraneal)0.976 x 1,147, (r² = 0,995). (6) COEFICIENTE DE ENCEFALIZACIÓN (EQ, en sus siglas en inglés, y así nombrada de aquí en adelante), que es un valor obtenido a partir de la relación alométrica del tamaño del cerebro con el tamaño del cuerpo. En este estudio se ha utilizado la fórmula establecida por Martin (1983) para el conjunto de los mamíferos placentados: EQ = masa cerebral / 11.22 x masa corporal0.76. (7) Dos variables de situación geográfica del yacimiento en el que aparecieron los restos: LATITUD y LONGITUD, expresadas en coordenadas geográficas. Se ha utilizado un mismo programa de sistema de posicionamiento geográfico (GoogleEarth) para obtener un mismo patrón localizador. La variable de latitud se ha planteado desde una perspectiva amplia de la zoogeografía, entendiendo unas latitudes bajas o tropicales (de 0 a 20 grados, sur o norte), zonas latitudinales semitropicales (de 20 a 40 grados, norte o sur) y zonas temperadas (a partir de 40 grados, norte o sur). La distribución geográfica de los individuos nos permitirá el estudio de los planteamientos ecogeográficos de Bergmann en el proceso de encefalización.
Tabla 1: Resumen de los datos utilizados en este estudio. GT: grupo taxonómico; ESP: especies incluidas; DAT: rango temporal de los fósiles, en kyr; N(VE): número de individuos para la variable de volumen endocraneal; N(MC): número de individuos para la masa corporal; N(EQ): número de individuos para el coeficiente de encefalización; N(TOT): número total de individuos. 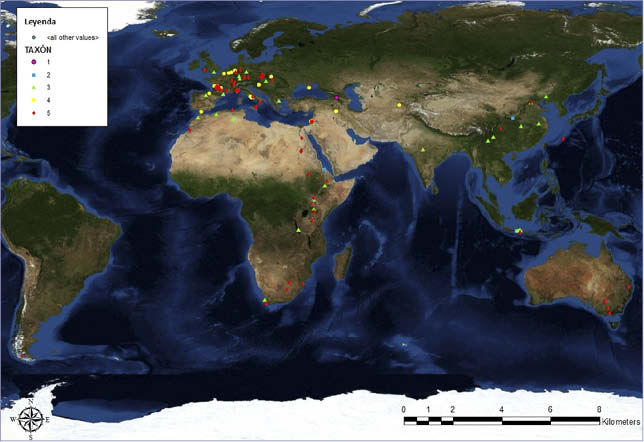 Figura 1: Mapa con la distribución de los individuos del Pleistoceno utilizados en este trabajo. Grupos taxonómicos: 1) habilinos; 2) erectinos del Pleistoceno inferior; 3) erectinos del Pleistoceno medio; 4) neandertales; 5) humanos anatómicamente modernos.
Se han considerado como variables principales el volumen endocraneal, la estimación de la masa corporal y el coeficiente de encefalización. Se han analizado dichas variables con métodos estadísticos descriptivos. Los valores considerados son el número de observaciones (N), el promedio ( Para contrastar la hipótesis de qué poblaciones son iguales o diferentes entre los distintos grupos, se ha utilizado un método para analizar la varianza, el ANOVA junto con el test post hoc de Bonferroni, el cual permite, una vez rechazada la hipótesis nula, controlando el nivel crítico, obtener una matriz que indica si las medias de los grupos son significativamente distintas entre sí. Para evaluar el grado de solapamiento entre los individuos de los diferentes grupos se ha utilizado la prueba de Lubischew. Previamente a su utilización, es necesario asegurarse que en nuestro conjunto de datos no se pueda rechazar la hipótesis de homocedasticidad y normalidad. Para el primer caso se ha aplicado el contraste de Levene, el cual analiza la igualdad o no de las varianzas de ambos grupos. Para contrastar la normalidad de los daros hemos utilizado el test de Shapiro-Wilk. Entonces pudimos aplicar el test de Lubischew, que está basado en la prueba de t-Student y permite evaluar el grado de solapamiento entre las distribuciones muestrales de dos poblaciones y, con ello, determinar qué capacidad tiene dicha variable de discriminar entre ambas poblaciones (LUBISCHEW 1962). El coeficiente de discriminación (K) se calcula a partir de la siguiente fórmula:
donde
cuanto mayor sea K mejor será el carácter X a la hora de discriminar. Así, cuanto mayor sea el valor de K, mejor será el carácter de la variable a la hora de discriminar. Se ha considerado utilizar el test de Lubischew porque mientras los análisis discriminantes incorporan la variabilidad del promedio con referencia a la media total, teniendo en cuenta la variabilidad tanto intergrupal como intragrupal, la prueba de Lubischew calcula la diferencia entre las medias de cada grupo, obteniendo unos resultados con una función de variación intergrupal. Por otro lado, el porcentaje de reclasificaciones correctas de la función discriminante es más sensible a la distribución de los datos dentro de cada grupo; mientras en el test de Lubischew las diferencias no dependen tanto de la distribución de los datos mismos, sino en la media, la desviación típica y el número de observaciones en el conjunto de los datos. Además la prueba de Lubischew es particularmente útil con muestras pequeñas, aunque como desventaja cabe resaltar que sólo se puede utilizar por parejas. Asimismo, el test de Lubischew ha sido utilizado en otros estudios para evaluar la discriminación entre taxones (PALMQVIST et al. 2007) y entre los sexos de una misma especie (JIMÉNEZ-ARENAS, 2010) con interesantes resultados. Para estudiar la influencia que algunas variables (por ejemplo, la latitud, el tiempo…) pudieran tener sobre el proceso de encefalización (representado por las variables principales: volumen endocraneal, masa corporal y coeficiente de encefalización) se han utilizado procedimientos de regresión y de correlación. La regresión permite descubrir la naturaleza de la relación entre dos variables usando una variable independiente para estimar los valores de una variable dependiente. A pesar de que la predicción es ciertamente difícil a partir de las ecuaciones de regresión, la utilización de éstas nos permite encontrar tendencias generales a partir de nuestros datos. La ecuación de regresión es aquélla que mejor se ajusta a los datos considerando que la distancia estimando la relación entre dos variables (x, y) además de la exactitud con la que los valores de y pueden derivarse de la estimación de x. La ecuación de regresión lineal es aquella que se ajusta mejor a los datos considerando que la distancia se toma en cada punto mediante las ordenadas y puede describirse así: y = a · xb siendo x la variable independiente e y la dependiente, a la ordenada en el origen y b la pendiente de la recta. Puede ajustarse a distintos tipos de gráficas (exponenciales, logarítmicas, polinomiales, lineales, etc.), aunque la más usual es el ajuste lineal. El método más usual de ajuste es el de los mínimos cuadrados. Por otro lado, la correlación mide la intensidad de la relación entre dos variables a través del coeficiente de correlación. Considerándolo en términos gráficos, el coeficiente de correlación es una medida del grado en el que los datos están dispersos alrededor de la recta de regresión, en el caso de que se trate de regresión lineal. Si los datos se encuentran muy próximos a dicha recta, la correlación será muy intensa y la predicción de y a partir de x será muy buena, y viceversa.
RESULTADOS
Tabla 2: Principales resultados de los estadísticos descriptivos (X: promedio; S: desviación típica; CV: coeficiente de variación) para las variables de VE, MC y EQ. El volumen endocraneal en los diferentes grupos taxonómicos En cuanto al volumen endocraneal (Tab. 2) a grandes rasgos se observa un incremento a lo largo del Pleistoceno. El grupo de los habilinos, con un promedio de volumen endocraneal de 635.36 cc, presentan valores que van desde 500 cc del Omo L 894-1 hasta los 777cc del individuo D-2280, siendo este primer grupo el que despliega una mayor heterogeneidad, puesto que ha proporcionado un valor mayor para el coeficiente de variación. El grupo de los erectinos del Pleistoceno inferior, con un promedio de 892.92 cc (con individuos que van desde los 779 cc del individuo de Gongwangling 1 hasta los 1069 cc del de OH9) se configuran como un grupo bastante homogéneo, presentando un valor del CV similar al que ofrecen los humanos anatómicamente modernos. La tendencia alcista en cuanto al tamaño del cerebro resulta aún más notable en el grupo de los erectinos del Pleistoceno medio (780-186 kyr). Este grupo, con un promedio de 1150.59 presenta una mayor heterogeneidad que el anterior, con volúmenes cerebrales que van desde los 727 cc del OH12 hasta otros muchos mayores como el KNM-ER 3884, con 1400 cc. Sin embargo, los mayores volúmenes endocraneales se encuentran en los individuos del Pleistoceno superior. El grupo de los neandertales, con un rango que abarca desde los 1065 cc de el de La Suard 1 a los 1745 cc del de Amud, con un promedio de 1425.4 cc. Los humanos anatómicamente modernos del Plesitoceno presentan un promedio algo mayor, 1462.23 cc, siendo un grupo más homogéneo que el de los neandertales, englobando valores que van desde los 1090 cc del individuo de Minatogawa 4 hasta los 1814 cc del de Barma Grande 2. A partir del Holoceno se constata una interrupción de esta tendencia creciente en cuanto al volumen endocraneal. El promedio para el grupo de los individuos del pueblo Pecos es de 1308.38 cc. Para rechazar la hipótesis nula de igualdad entre las medias del volumen endocraneal entre los diferentes grupos aplicamos el test post hoc de Bonferroni, el cual da como resultado que todas las medias del tamaño del cerebro son significativamente distintas (a<0.05) entre sí dos a dos, excepto las de los neandertales y los humanos anatómicamente modernos del Pleistoceno. Los valores del test de Shapiro-Wilk nos permiten descartar que los datos no presentan una distribución normal (a>0.05). La prueba de Levene (s=0.037) confirma que la varianza no es homogénea en los grupos, aunque tampoco se desvía demasiado del valor crítico (a= 0.05). En los resultados del test de Lubischew para el volumen endocraneal (Tab. 3) se observa que, durante el Pleistoceno, el grado de solapamiento entre los grupos va en aumento, hasta alcanzar el máximo en la pareja neandertal – humanos anatómicamente modernos del Pleistoceno, donde se ha obtenido un porcentaje de solapamiento del 45.22% (Fig. 2). Por otro lado, el grado de solapamiento es mayor entre neandertales y Pecos que entre éstos últimos y los humanos anatómicamente modernos del Pleistoceno (34.83% frente a 28.77%). 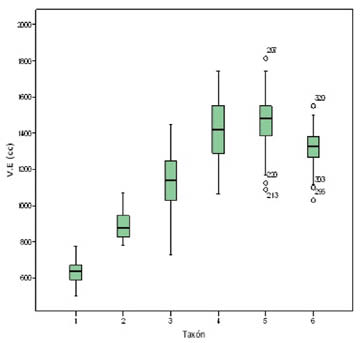 Figura 2: Diagrama de cajas del volumen endocraneal de los distintos grupos taxonómicos.
Tabla 3: Resultados del test de Lubischew. Donde N es el número de observaciones; R es el coeficiente de determinación y se ofrec eun porcentaje de grado de solapamiento y otro de la capacidad discriminante de cada variable (volumen endocraneal, masa corporal y coeficiente de encefalización) entre los pares de taxones.
La estimación de la masa corporal en los distintos grupos taxonómicos El escaso número de estimaciones de la masa corporal de los dos primeros grupos (Tab.2), habilinos y erectinos del Pleistoceno inferior, ha imposibilitado su uso para los análisis estadísticos. La masa corporal también aumenta a los largo del Pleistoceno. Los erectinos del Pleistoceno medio presentan un promedio de 66.56 kg, con un importante grado de heterogeneidad, como ejemplifican los casos del individuo fósil de Gesher Benot Ya’akov (53.4 kg) y el de Boxgrove (86.7 kg). El aumento de la masa corporal es aún más notable en el grupo de los neandertales, con un promedio de 72.93 kg y el menor coeficiente de variación, 10.46% para esta variable. A partir de los humanos anatómicamente modernos el promedio de la masa corporal se ve reducido: 64.11 kg para los del Pleistoceno y 55.51 kg para los del Holoceno. El test post hoc de Bonferroni no nos permitió rechazar la hipótesis nula de igualdad entre los promedio de la masa corporal entre algunos de los grupos considerados. Si que resultaron significativamente distintas (a<0.05) las de neandertales con los erectinos del Pleistoceno inferior y con los humanos anatómicamente modernos, tanto del Pleistoceno como del Holoceno. Los valores de Shapiro-Wilks (a>0.05) nos permite descartar que los datos no presenten una distribución normal. La prueba de Levenne (s=0.018) confirma que la varianza no es homogénea entre los grupos, aunque, nuevamente, no se aparta en exceso del valor crítico. Como los promedios de la masa corporal de neandertales y humanos anatómicamente modernos (tanto del Pleistoceno como del Holoceno) sí son significativamente distintos entre sí, aplicamos el test de Lubischew para estudiar el grado de solapamiento en esta variable (Tab. 3). Los resultados nos muestran un porcentaje de solapamiento muy bajo entre los tres grupos, sobre todo en el caso de la masa corporal entre neandertales y humanos anatómicamente modernos del Pleistoceno (10.77% de solapamiento), lo que significa que esta variable es altamente discriminante entre ambos grupos. El coeficiente de encefalización (EQ) en el género Homo Para los grupos taxonómicos más antiguos (habilinos y erectinos) no hay individuos fósiles que nos permitan estimar a la vez el volumen endocraneal y la masa corporal por lo que no se ha podido calcular el coeficiente de encefalización (Tab. 2). En el grupo de los erectinos del Pleistoceno medio tan sólo consta de un individuo, el de Jinnu Shan, que alcanza un valor de 4.145 (obviamente no se ha utilizado en los análisis estadísticos). El promedio que se ha obtenido para el grupo de los neandertales es de 4.99 frente a 5.43 de los humanos anatómicamente modernos. Además estos últimos se caracterizan por una fuerte homogeneización, con un coeficiente de variación de 6.84. Al aplicar el t-student entre ambos grupos se obtuvo que sí existen diferencias significativas entre los promedios de ambos grupos (T=3.124; s=0.03). Esto fue corroborado por el post hoc de Bonferroni, para ésta pareja, pero en ningún caso entre estos grupos con los humanos anatómicamente modernos del Holoceno. Los valores de Saphiro-Wilk (a>0.05) nos permite descartar que los datos no presentan una distribución normal. La prueba de Levene (s=0.009), muy ajustada al nivel crítico. Al aplicar el test de Lubischew (Tab. 3), resulta interesante que los neandertales presentan un mayor grado de solapamiento con los humanos anatómicamente modernos del Holoceno (37.04%) que con los del Holoceno (28.31%). El solapamiento entre los humanos modernos es muy alto (46.18%). Encefalización y ecogeografía A grandes rasgos, los grupos taxonómicos más antiguos se concentran en latitudes bajas (de 0 a 20 grados) a pesar de que presentan un grado de heterogeneidad altísimo. En el Pleistoceno medio, la dispersión geográfica se multiplica, hallándose los fósiles en todas las zonas latitudinales, con un promedio de unos 25.73º. Los neandertales resultan el grupo más homogéneo, con un promedio de 42.96º, restringiéndose sus hallazgos al continente euroasiático. Los humanos anatómicamente modernos son los grandes colonizadores del planeta, y ocupan la totalidad de las franjas latitudinales, amén de las longitudinales. Su promedio es de 36.18º. A partir de los análisis de regresión (Tab. 4), tanto al conjunto de los taxones (Homo) como en los diferentes grupos, no se han hallado grandes expresiones, por lo general, de relación entre la latitud como variable independiente, y las principales variables manejadas en el presente estudio. El volumen endocraneal no presenta relación con la latitud para cada uno de los grupos, aunque tomando en conjunto al género Homo si existe una relación estadísticamente significativa entre el volumen endocraneal y la latitud, a mayor latitud, mayor volumen endocraneal. En cuanto a la masa corporal ocurre algo similar, existe una relación, esta vez algo más débil, entre la masa corporal para el género Homo y la latitud, al igual que considerando los humanos anatómicamente modernos, a mayor latitud, mayor masa corporal, concordando con la regla ecogeográfica de Bergman. Para el coeficiente de encefalización y la latitud no se han obtenido relaciones estadísticamente significativas, aunque los neandertales muestran una tendencia a poseer menores coeficientes de encefalización a mayor latitud, siendo posible que el escaso número de observaciones para el grupo de los neandertales imposibilite la significación estadística.
Tabla 4: Resultados del coeficiente de determinación de Pearson (R) y su significación (s) entre la latitud (como variable independiente) y el volumen endocraneal, la masa corporal y el coeficiente de encefalización. Tiempo y encefalización Al aplicar análisis de regresión (Tab. 5) entre el tiempo (como variable independiente) y el volumen endocraneal para el conjunto del género Homo se halló una relación fuerte entre ambas (R=-0.829, s=0.000). El carácter negativo de la relación viene dado por el ordenamiento descendente de los datos, desde lo más antiguo a lo más reciente. Para los diferentes taxones, la única relación estadísticamente significativa se ha dado en el grupo de los neandertales, tanto con el volumen endocraneal como para la masa corporal. Esto significa que aquellos neandertales más recientes presentan mayores tamaños cerebrales y corporales.
Tabla 5 :Resultados del coeficiente de determinación de Pearson (R) y su significación (s) entre el tiempo (como variable independiente) y el volumen endocraneal, la masa corporal y el coeficiente de encefalización.
El volumen endocraneal El claro aumento del tamaño del cerebro durante la evolución del género Homo está aceptado por la mayoría de autores, aunque surgen discrepancias a la hora de valorar dicho proceso. En ocasiones, las pequeñas discordancias vienen marcadas por la propia naturaleza de la muestra, sesgada y con un número de observaciones muy diferentes para cada uno de los taxones, o bien por las diversas clasificaciones taxonómicas utilizadas por cada autor. Algunos autores apuntan a que el mayor incremento del volumen endocraneal se dio en el Pleistoceno Medio (600 – 150 kyr), y que dicho momento fue precedido por un largo periodo de stasis (RUFF et al. 1997). Sin embargo, Holloway (1999) apunta a que no se puede hablar de stasis durante el proceso de encefalización del género Homo, porque, como se ha visto anteriormente, hay que tener en cuenta las reorganizaciones del espacio cerebral, que podrían haber tenido lugar sin necesariamente implicar un aumento del tamaño del encéfalo. Por otro lado, Ruff et al. (1997), ponen de manifiesto que el incremento perceptible en el Pleistoceno superior, uno de los factores aludidos por estos autores podría ser el hallazgo de fósiles en latitudes más altas, por lo que lo vinculan con la regla ecomorfológica de Bergmann que vincula el aumento del tamaño con el clima y la latitud. Por otro lado, tal aumento podría estar relacionado también con la variabilidad en el tamaño del cerebro entre el género masculino y femenino, donde las mujeres presentan, por lo general, menores volúmenes endocraneales que los hombres, debido a que, de igual forma, presentan tamaños corporales, generalmente, más pequeños. No obstante, es bien conocido que el dimorfismo sexual en el volumen endocraneal es reducido incluso para aquellas especies que presentan un notable dimorfismo sexual en el tamaño corporal [vg. gorilas y orangutanes (JIMÉNEZ-ARENAS et al., 2006)]. El porcentaje de solapamiento en cuanto al volumen endocraneal indica que dicha variable presenta una razonable fiabilidad para distinguir taxones. Una interesante excepción ocurre con el caso de neandertales y humanos anatómicamente modernos del Pleistoceno. Por tanto, el cambio en el volumen endocraneal implica un componente taxonómico, pero también temporal, puesto que los erectinos se dividen en dos grupos claramente diferenciados: los del Pleistoceno inferior y los del Pleistoceno medio, lo cual es consistente con el estudio previo de Rightmire (2004), el cual observa, a partir de las diferencias en el volumen endocraneal y de otras características craneofaciales, una distinción entre ambos grupos de erectinos que el interpreta en términos taxonómicos (identificando una cronoespecie que podría adscribirse a Homo heidelbergensis). Sin embargo, nosotros interpretamos dicha variación dentro del grupo de los erectinos como consecuencia de un cambio temporal en vez de taxonómico (JIMÉNEZ-ARENAS et al., 2011). Al respecto de los individuos del Pleistoceno superior, el promedio del volumen endocraneal es ligeramente superior en los humanos anatómicamente modernos que en los neandertales. Tradicionalmente (STRINGER, 1992; HOLLIDAY, 2008), se ha considerado que los neandertales poseían un cerebro mayor en términos absolutos, sin embargo, tal afirmación puede estar relacionado con la muestra utilizada. Si en la muestra de humanos anatómicamente modernos se incluyen tanto los del Pleistoceno como los del Holoceno, entonces el promedio para el conjunto de Homo sapiens es ligeramente inferior al de los neandertales (SERRANO-RAMOS et al., 2011), debido a que los humanos modernos actuales poseemos un volumen endocraneal menor que nuestros ancestros del Pleistoceno. Dicho decrecimiento en el tamaño del cerebro también va acompañado de una disminución del tamaño corporal. En nuestro caso de estudio, el grupo que representa a los humanos del Holoceno, los Pecos, pueden presentar una disminución de la masa corporal que puede estar también relacionada con aspectos biológicos / culturales propios de este pueblo, por lo que se precisaría un estudio enfocado a cuantificar dicho decrecimiento en el tamaño del cerebro/cuerpo a partir del Holoceno. Sin embargo, no parece posible explicar la disminución del encéfalo sólo en relación al decrecimiento de la masa corporal. Al respecto, recientes investigaciones en genética han mostrado la existencia de dos genes, el MCPH 1 y ASPM, relacionados con la disminución del tamaño del cerebro y que podrían explicar parte de esta reducción, teniendo el cuenta que la presencia del MCPH 1, un gen que regula el tamaño del cerebro y que ha evolucionado bajo una fuerte selección positiva en el linaje de los humanos anatómicamente modernos y cuyo efecto está constatado a partir del 37000 BP (EVANS, 2004; 2005), y la del ASPM, un regulador específico del tamaño del cerebro y cuya evolución dentro del mismo linaje tuvo una fuerte selección positiva a partir del 5800 BP (MEKEL-BROBOV, 2007). Por otro lado, resultan interesantes ciertos estudios que relacionan el incremento de la densidad de población, con un generalizado, aunque ligero, decrecimiento del volumen endocraneal. Esto podría haber coincidido con la emergencia de la agricultura y pueblos y ciudades-estado más grandes y social y económicamente organizados (BRACE, 1995), lo cual estaría más próximo a su relación con la evolución del ASPM que con la del MCPH1. Lo cierto es que, podemos entender fácilmente que tanto los cambios acaecidos a partir del Holoceno, tanto a nivel climático, geográfico, ecológico y sobre todo, social y cultural, permite separar a nuestros ancestros pleistocenos de sus descendientes holocenos, a pesar de ser parte de la misma especie. No obstante, como hemos visto, hay numerosos investigadores que abogan por la preponderancia de lo social sobre lo natural en la evolución humana desde tiempos tempranos (ALEXANDER; 1989; HOLLOWAY, 1975; DURBAN, 1998; BAILEY & GEARY, 2009). Sin embargo, nosotros optamos por considerar como fundamentales las interacciones que se producen entre los diferentes niveles considerados. Volviendo al tamaño absoluto del cerebro, menor, de los neandertales que en los humanos anatómicamente modernos, remarcar, sin embargo, que de esta conclusión no derivamos la inferioridad cognitiva de los neandertales, pues está sobradamente demostrada (aunque no por todos aceptada) su capacidad, tanto a nivel de supervivencia como a nivel simbólico, por ejemplo a través de los enterramientos o de adornos personales y uso de pigmentos (ZILHÃO, 2010). Ecogeografía y encefalización En este estudio hemos pretendido cuantificar la dispersión geográfica de los individuos utilizados y tratar de analizar la posible relación con ciertos principios ecogeográficos y el proceso de encefalización. Las reglas ecogeográficas que relacionan la morfología del cuerpo con el clima, como la de Bergmann (1847) o la de Allen (1877), son casos específicos de una relación teórica más general entre la masa corporal, la superficie del cuerpo y la temperatura ambiente (MAYR, 1963; SCHREIDER, 1964; ROBERTS, 1978; RUFF, 1991). En una especie ampliamente distribuida, el ratio entre la superficie corporal y la masa corporal se incrementaría en los climas cálidos y decrecería en climas más fríos, para promover o retardar la pérdida de calor, respectivamente. Existen numerosas evidencias empíricas que sustentan estas teorías, tanto en humanos como en animales (RENSCH, 1936, 1960; SCHREIDER, 1950; ROBERTS, 1953; NEWMAN 1953; MAYR, 1963; TRINKAUS, 1981; RUFF, 1991). A pesar de todo, la validez de estos presupuestos ecogeográficos, especialmente sus aplicaciones en humanos, ha sido cuestionada por diversos investigadores (SCHOLANDER, 1955; WILBER, 1957; MCNAB, 1971). Las críticas a la influencia del clima /variabilidad latitudinal en la morfología corporal se basan sobre todo en que, dichas reglas no se cumplen en todas las especies [aunque sí en la mayoría de especies politípicas con un amplio rango de distribución geográfica (RUFF, 1991)], en la importancia de otros factores (v.g. la disponibilidad de recursos alimenticios en todas las zonas ocupadas), o mecanismos fisiológicos que influyan en la termorregulación. Para el caso de los humanos, las principales discrepancias al respecto de la variabilidad en la masa corporal por efectos del clima vienen marcadas por el papel de la cultura versus medioambiente, es decir, que en los humanos los presupuestos ecogeográficos (en concreto en referencia a condiciones climáticas frías) se habrían mitigado con la cultura (uso de refugios, ropajes y pieles, control del fuego). Con todo, en los humanos modernos actuales está demostrada la influencia de los patrones ecogeográficos (RUFF, 1991), con lo que, con más razón, es posible que afectara a las poblaciones extintas de Homo que, asimismo, están menos afectados por la cultura. Al aplicar estudios de regresión para analizar la posible relación entre la latitud y el volumen endocraneal, se ha hallado una relación moderada para el conjunto de los taxones y la latitud. En conjunto, se puede considerar que el volumen endocraneal para el género Homo responde a los principios ecogeográficos de Bergmann, de forma que, el tamaño del cerebro se incrementa, moderadamente, al alejarse las poblaciones del Ecuador. Asimismo, se puede hacer una inferencia similar para los humanos modernos pleistocenos. En cuanto a la relación entre la masa corporal y la latitud, ésta vuelve a ser, de nuevo, significativa estadísticamente para el conjunto del género Homo y para los humanos anatómicamente modernos del Pleistoceno, aunque en ambos casos la relación es de carácter débil. Esto concordaría con los principios ecogeográficos de Bergmann, que predice que el tamaño corporal se incrementa conforma lo hace la latitud. A modo de conclusión Se da un evidente incremento en el volumen endocraneal en el género Homo, en clara relación en su conjunto con cuestiones temporales y taxonómicas. En el proceso evolutivo se pueden identificar dos grandes momentos donde el incremento del volumen endocraneal es más reseñable que en el resto de los grupos taxonómicos: los erectinos del Pleistoceno inferior y los humanos anatómicamente modernos del Pleistoceno. A tenor de los resultados aquí obtenidos, el volumen endocraneal supone una variable de razonable fiabilidad para distinguir grupos taxonómicos, a excepción de entre neandertales y humanos anatómicamente modernos. Además, el promedio del tamaño absoluto del cerebro de los neandertales es ligeramente inferior al de los humanos anatómicamente modernos del Pleistoceno, en contra de lo propuesto por algunos autores y de lo arraigado de este supuesto en la ciencia divulgativa. La variabilidad en el tamaño del cerebro observada tanto en neandertales como en los humanos anatómicamente modernos parece responder a tendencias diferenciadas. Mientras los humanos anatómicamente modernos encajan con los presupuestos ecogeográficos de Bergmann, presentando una relación entre la masa corporal y la latitud en que se hallaron los fósiles; los neandertales, por su lado, responden más bien a cuestiones temporales, por las cuales, tanto el cerebro como el tamaño corporal en general es mayor en aquellos neandertales más modernos. Esto podría relacionarse con un impedimento evolutivo por el cual el tamaño del cerebro de los neandertales se incremente a la par que lo hace el tamaño corporal de éstos. Por otro lado, la restringida dispersión geográfica de los neandertales (prácticamente confinados en el continente euroasiático) impida que estás poblaciones se vean afectadas por los patrones ecogeográficos. Con todo, no parece que los principios ecogeográficos de Bergmann puedan ser universales para todos los individuos del género Homo, ya que, aunque considerando en conjunto a Homo sí se han hallado relaciones estadísticamente significativas que son consistentes con estas reglas ecogeográficas, parece que analizando los distintos grupos taxonómicos (a excepción de los humanos anatómicamente modernos del Pleistoceno) la relación con los presupuestos ecogeográficos no está tan clara.
La autora agradece al Dr. C.B. Ruff, el cual me cedió sus datos generosamente y tuvo a bien hacerme llegar las observaciones oportunas. Y al Dr. J.M. Jiménez Arenas y J.A. Esquivel Guerrero por su dedicación y en la dirección de este trabajo.
AIELLO, L.C. & WOOD, B. (1994): Cranial variables as predictors of hominine body mass. Current Anthropology 34: 184-193. AIELLO, L.C. & WHEELER, P. (1995): The expensive-tissue hypothesis: the brain and the digestive system in human and primate evolution. Current Anthropoly 36: 199-221. ALEXANDER, R. D. (1989): Evolution of the human psyche. In: MELLARS, P. & STRINGER, C. (Eds.) The human revolution: Behavioural and biological perspectives on the origins of modern humans. (pp. 455-513). Princeton University Press, Princenton New Jersey, 1989. ALLEN, J.A. (1877): The influence of physical conditions on the genesis of species. Radical Rev. 1:108-140. ASH, J. & GALLUP, G.G. (2007): Brain size, intelligence, and paleoclimatic variation. In: GEHER, G. & MILLER, G. (eds.). Mating Intelligence: Sex, Relationships and the Mind’s Reproductive System. Lawrence Erlbaum Associates, New York, 2007, pp 311-333 BAILEY, D.H. & GEARY, D.C. (2009): Hominid brain evolution. Testing climatic, ecological and competition models. Human Nature 20: 67-79. BARTON, R.A. (1996): Neocortex size and behavioural ecology in primates. Proceedings of the Royal Biological Sciences Society 263, 173-177. BERGMANN, C. (1847): Uber die verhaltniesse der warmeokonomie der thiere zu ihrer grosse. Göttingen Stud 1: 595-708. BRACE, C.L. (1995): Biocultural interaction and the mechanism of mosaic evolution in the emergente of “modern” morphology. American Anthropologist 97:711-721. DE MIGUEL, C. & HENNERBERG, M. (2001): Variation in hominid brain size: How much is due to the method? Homo 52: 3-58. DURBAN, R.I.M. (1998): The social brain hypothesis. Evolutionary Anthropology 6: 178 – 190. EVANS, P.D., ANDERSON, J.R., VALLENDER, E.J., CHOI, S.S., LAHN, B.T.(2004): Reconstructing the evolutionary history of Microcephalin, a gene controlling human brain size. Human Molecular Genetics 13: 1139-1145. EVANS, P.D., GILBERT, S.L., MEKEL-BOKOROV, N., VALLENDER, E.J., ANDERSON, J.R., VAEZ-AZIZI, L.M., TISHKOFF, S.A., HUDSON, R.R. & LAHN, B.T. (2005) Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans. Science 309: 1717-1145. HOLLIDAY, T.W (2008). Neanderthals and modern human: an example of a mammalian syngameon? In. Neanderthal Revisited: New Approaches and Perspectives (eds. Hublin, J.J., Havarti, K & Harrison). Springer, version digital, 2008, pp: 281-298 HOLLOWAY, R.L. (1967): The evolution of the human brain: Some notes towards a synthesis between neural structure and the evolution of complex behaviour. General Systems 12: 3-19. HOLLOWAY, R.L. (1975): The Role of Human Behaviour in the Evolution of the Brain. [The 43rd James Arthur Lecture on the evolution of the human brain at the American Museum of Natural History, 1973]. The American Museum of Natural History, New York, 1973. HOLLOWAY, R.L. (1999): Evolution of the Human Brain. In: Handbook of Human Symbolic Evolution. (eds. Lock, A. & Peters, C.). Blackwell Publisher, Ltd, 1999, Oxford. HOLLOWAY, R.L. (2009): Brain Fossils: Endocasts. In: Encyclopedia of Neuroscience. (ed. Squire, L.R.), volume 2. Academic Press, 2009, Oxford, pp: 353-361. JERISON, H.J. (1973): Evolution of the brain and intelligence. Academic Press, 1973, New York. JIMÉNEZ ARENAS, J.M. (2006): Estudio de la Variación Craneométrica en Homínidos y sus Inferencias Paleobiológicas y Filogeenéticas. Tesis Doctoral. Universidad de Granada, 2006, Granada. JIMÉNEZ ARENAS, J.M. (2010): Discriminación de Sexo en una Población Medieval del Sur de la Península Ibérica Mediante el Uso de Variables Simples. Internacional Journal of Morphology, 28(3): 667-672. JIMÉNEZ ARENAS, J.M., PÉREZ-CLAROS, J.A. & PALMQVIST, P. (2011) A probabilistic approach to the craniometric variability of the genus Homo and inferences on the taxonomic affinities of the first human population dispersing out of Africa. Quaternary International 243-1: 219-230. KAPLAN, H., HILL, K., LANCASTER, J., & HURTADO, A.M (2000): A theory of human life history evolution: diet, intelligence and longevity. Evolutionary Anthropology 9: 156-185. LEE, A. & PEARSON, K. (1901): Data for the Problem of Evolution in Man. VI. A First Study of the Correlation of the Human Skull. Philosophical Transactions of the Royal Society. Pp: 225-264. LOW, B.S. (1990): Marriage systems and pathogen stress in human societies. American Zoologist 30: 325-339. LUBISCHEW, A. A. (1962): On the use of discriminant functions in taxonomy. Biometrics 18:455-477. MARTIN, R.D. (1983): Human Evolution in an Ecological Context. Fifty-second James Arthur Lecture. American Museum of Natural History, 1983, New York. MATARESE, G. & LA CAVA, A. (2004): The intrincate interface between inmune system and metabolism. Trends in Immunology 25: 193-200. MAYR, E. (1963): Animal Species and Evolution. Harvard University Press, 1963 Cambridge. MEKEL-BOBROV, N., POSTHUMA, D., GILBERT, S.L., LIND, P., GOSSO, M.F., LUCIANO, M., HARRIS, S.E., BATES, T.C., POLDERMAN, T.J.C, WHALLEY, L.J., FOX, H., STARR, J.M., EVANS, P.D., MONTGOMERY, G.W., FERNANDES, C., HEUTINK, P., MARTIN, N.G., BOOMSMA, D.I., DEARY, I.J., WRIGHT, M.J., DE GEUS, E.J.C. & LAHN, B.T. (2007): The ongoing adaptative evolution of ASPM and Microcephalin is not explain by increased intelligence. Human Molecular Genetics 16: 600-608. MCNAB, B.K. (1971): On the ecological significance of Bergmann’s rule. Ecology 52: 845-854. MILTON, K. (1981): Distribution patterns of tropical plant foods as an evolutionary stimulus to primate metal development. American Anthropologist 83: 534-548. MILTON, K. (1983): Diet and primate evolution. Scientific American 269: 86-93. NEWMAN, M.T. (1953): The applications of ecological rules to the racial anthropology of the aboriginial new world. American Anthropology 55: 311-327. PALMQVIST, P., TORREGROSSA, V., PÉREZ-CLAROS, J.A., MARTÍNEZ-NAVARRO, B. & TURNER, A. (2007): A re-evaluation of the diversity of Megantereon (Mammalia, Carnivora, Machairodontinae) and the problem of species identification in extinct carnivores. Journal of Vertebrate Paleontology 27: 160-175. POTTS, R. (2001): Variability selection in hominid evolution. Evolutionary Anthropology 7: 81-96. RENSCH, B. (1936): Studien uber klimatische paralleitat der merkmalaupragung bei vogeln und saugern. Archiv für Naturgeschichte 5: 317-363. RENSCH, B. (1960): Evolution Above the Species Level. Columbia University Press, 1960, New York. RIGHTMIRE, G.P. (2004): Brain size and encephalization in early to mid Pleistocene Homo. American Journal of Physical Anthropology 12: 109-123. ROBERTS, D.F. (1953): Body weight, race and culture. American Journal of Physical Anthropology 11: 533-558. ROBERTS, D.F. (1978): Climate and Human Variability. (2nd edn). Cummings, 1978, Menlo Park, California. RUFF, C.B. (1991): Climate and body shape in hominid evolution. Journal of Human Evolution 21: 81-105. RUFF, C.B. (1994): Morphological adaptation to climate in modern and fossil hominids. Yearbook of Physical Anthropology 37: 65-107. RUFF, C.B., TRINKAUS, E. & HOLLIDAY, T.W. (1997): Body mass and encephalization in Pleistocene Homo. Nature 387: 173-176. SCHOLANDER, P.F. (1955): Evolution of climatic adaptation in homeotherms. Evolution 9: 15-26. SCHREIDER, E. (1950): geographical distribution of the body-weight/body-surface ratio. Nature London 165: 286. SCHREIDER, E. (1964): Ecological rules, body-heat regulation, and human evolution. Evolution 18: 1-9. SERRANO RAMOS, A., JIMÉNEZ ARENAS, J.M. & ESQUIVEL GUERRERO, J.A. (2011): Volumen endocraneal en Homo: implicaciones taxonómicas. En: XXXVII Jornadas de la Sociedad Española de Paleontología. Simposios de los proyectos PICG 587 y 596. Paleontologia i Evolució. Institut Català de Paleontologia Miquel Crusafont, 2011, Sabadell. STRINGER,C.B.(1992). Evolution of early humans. In: The Cambridge Encyclopedia of Human Evolution. (eds. JONES, S., MARTIN, R. & PILBEAM, D.) Cambridge University Press, 1992, Cambridge. THOMPSON, D.W. (1917) On Growth and form. Cambrigde University Press, 1917, Cambridge. TRINKAUS, E. (1981): Neanderthal limb proportions and cold adaptation. In (STRINGER, C.B. Ed.) Aspects of Human Evolution. Taylor & Francis, 1981, London, pp: 187-224. WILBER, C.G. (1957): Physiological regulations and the origin of human types. Human Biology 29: 329-336. WOOD, B. (2005): Human evolution: a brief insight. Sterling Publishing Co., 2005, New York. ZILHÃO, J., ANGELUCCI, D.E., BADAL-GARCÍA, E., D’ERRICO, F., DANIEL, F., DAYET, L., DOUKA, K., HIGHAM, T.F.G., MARTÍNEZ-SÁNCHEZ, M.J., MONTES-BERNÁRDEZ, R.J., MURCIA-MASCARÓS, S., PÉREZ-SRIVENT, C., ROLDÁN-GARCÍA, C., VANHAEREN, M., VILLAVERDE, V., WOOD, R. & ZAPATA, J. (2010): Symbolic use of marine shells and mineral pigments by Iberian Neanderthals. Proceedings of the National Academy of Science 107 (3): 1023-1028. * Departamento de Prehistoria y Arqueología, Universidad de Granada. alisera@correo.ugr.es. |