
APROXIMACIÓN ARQUEOZOOLÓGICA A LA EDAD DEL BRONCE EN EL SURESTE PENINSULAR: EL YACIMIENTO DE CUESTA DEL NEGRO (PURULLENA, GRANADA). UN ESTUDIO TAXONÓMICO Y TAFONÓMICO
ARCHAEOZOOLOGICAL APPROACH TO THE BRONZE AGE IN PENINSULA’S SOUTHEAST: CUESTA DEL NEGRO SITE (PURULLENA, GRANADA). A TAXONOMIC AND TAPHONOMIC STUDY
Lucía TINOCO DOMÍNGUEZ *
Resumen
Este trabajo presenta un análisis del aprovechamiento y consumo de las diferentes especies animales de La Cuesta del Negro (Purullena, Granada). A partir del estudio taxonómico y tafonómico de los restos arqueozoológicos de este yacimiento hemos extraído algunas conclusiones sobre los modos de explotación faunística tanto en el área superior como en la inferior de este asentamiento. El presente artículo aprovecha los resultados obtenidos para apreciar la evolución de dichas prácticas en las distintas fases de este periodo cronológico.
Palabras clave
Cuesta del Negro. Arqueozoología. Taxonomía. Tafonomía. Edad del Bronce.
Abstract
This study presents an analysis of the usage and consumption of different animal species of the Bronze Age Cuesta del Negro site (Purullena, Granada). By performing taxonomic and taphonomic observations of the faunal remains found in this area conclusions can be drawn to determine the peculiarities pertaining the nature of the exploitation of the species at stake in both the upper and lower areas of this settlement. This paper uses the results obtained to outline the evolution of this phenomenon through the different phases of this time period.
Key Words
Cuesta del Negro. Archaeozoology. Taxonomy. Taphonomy. BronzeAge.
INTRODUCCIÓN
La relativa escasez de publicaciones sobre la Edad de Bronce Tardío y Final (c. 1550-750/700 cal AC) (JOVER MAESTRE et al. 2016:Fig. 3) en el sureste peninsular hasta la fecha es consecuencia tanto del menor interés de los investigadores por las evidencias conservadas para este periodo como por la aparente escasez de las mismas. Sea como fuere, durante este periodo final se fueron perdiendo las prácticas argáricas de forma paulatina: los asentamientos fueron abandonados o su urbanismo fue transformado, muchos útiles típicamente argáricos cayeron en el olvido, las costumbres funerarias argáricas dejaron de ser practicadas, etc. Todo ello se tradujo como una ruptura a nivel económico y social (ARANDA JIMÉNEZ 2015:137). Es en este proceso de cambio en el que puede encuadrarse la Cuesta del Negro (Purullena, Granada) y en el que gana importancia al permitir evaluar dichas transformaciones en un mismo lugar, habitado tanto en el Bronce Antiguo como en el Bronce Tardío. En esta ocasión, hemos decidido acercarnos a esta cuestión a partir de un estudio taxonómico y tafonómico de restos arqueozoológicos procedentes de los dos periodos cronológicos de la Cuesta del Negro y de sus dos áreas de ocupación, que posteriormente desarrollaremos. De esta forma, pretendemos establecer una comparativa cronológica y espacial sobre los modos de explotación faunística desde el Argar hasta el Bronce Tardío para apreciar variaciones en la preferencias de unas especies a otras, en la elección de individuos de determinada edad o sexo, posible aprovechamiento de los mismos para la obtención de productos secundarios o de su fuerza de tiro o carga, etc.
La primera ocupación de este enclave se produjo en el Bronce Antiguo por poblaciones argáricas, que se mantuvieron en este lugar hasta el Bronce Pleno. Los análisis radiocarbónicos han ofrecido unas fechas entre el 1850 y el 1600 a.C. para este periodo. Tras un breve hiato de tiempo el asentamiento volvió a ser habitado en el Bronce Tardío por personas adscritas a la cultura mesteña de Cogotas I, en concreto entre el 1550 y el 1350 a.C. según las fechas radiocarbónicas (CONTRERAS CORTÉS 1986:278). Esta cultura arqueológica ha sido definida a partir de la presencia de decoraciones cerámicas a boquique o a bandas incisas, excisas o impresas (CASTRO MARTÍNEZ et al. 1995:51).
A nivel espacial, la Cuesta del Negro se localiza en el término municipal de Purullena (Granada) (Fig. 1), integrado en la Depresión de Guadix, en concreto, en una cuesta de 500 m de altitud compuesta por conglomerados, arenas y lutitas que producen unos relieves tipo bad-lands (MOLINA GONZÁLEZ y PAREJA LÓPEZ 1975:9-12; SÁEZ PÉREZ y ARRIBAS PALAU 1979:22). Se trata de un lugar estratégico protegido por barrancos y otros accidentes geográficos. Seguramente, su altura diferenciada permitió a las personas que la habitaron controlar visualmente las rutas de tránsito entre el oeste y el sur peninsular por la Depresión Penibética (PÉREZ IBÁÑEZ 2011:68-69), así como mantener bajo observación al río Fardes que discurre muy próximo al poblado. Esta población se abastecería de esta corriente hídrica y aprovecharía las tierras regadas por la misma para el desarrollo de la agricultura. De igual modo, este enclave contaba con pastos adecuados en su entorno para el buen desarrollo de la ganadería.
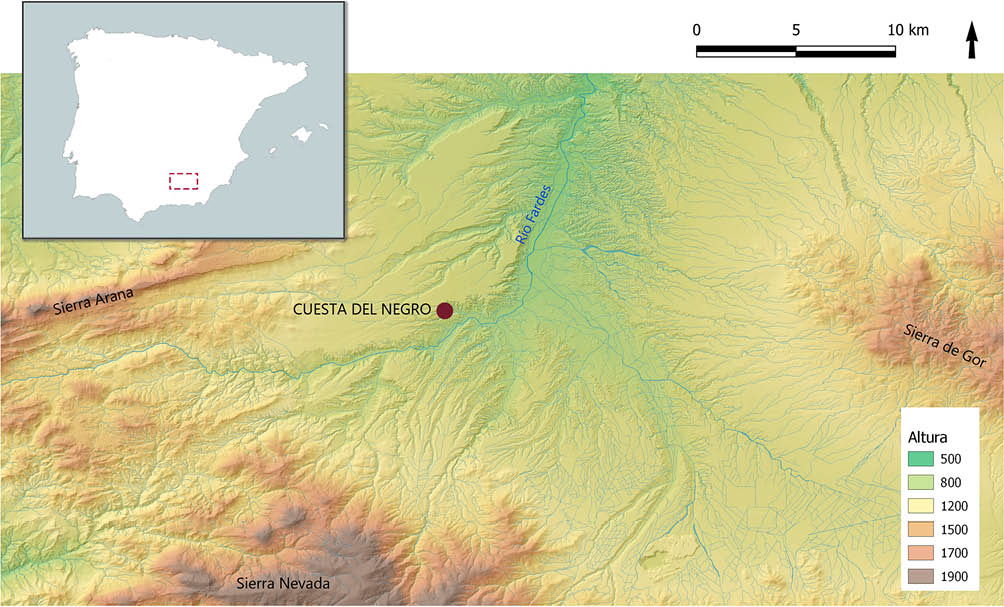
Fig. 1. Ubicación del yacimiento de la Cuesta del Negro (Purullena, Granada) a nivel peninsular y dentro de su entorno geográfico. Elaboración propia.
La Cuesta del Negro se divide internamente en dos áreas, una inferior y otra superior; distribución que responde a las características del modelo descrito para el Grupo Granadino de la Cultura de El Argar (MOLINA GONZÁLEZ y CÁMARA SERRANO 2009:199-200). El área inferior se sitúa en la pendiente del cerro y es allí donde se desarrolló el hábitat per se. Esta área presenta viviendas compartimentadas con planta de tendencia rectangular y con sepulturas individuales bajo sus suelos para el periodo argárico (Ibídem: 203). En cambio, durante el periodo de Cogotas I se produce una reorganización de las estructuras de hábitat en torno a un eje central. Las mismas muestran una planta rectangular más regular, suelos de barro endurecido y un hogar en su zona central (CONTRERAS CORTÉS 1986:287).
El área superior se corresponde con un sistema fortificado que se distribuye entre dos cerros y que está conformado por un fortín de planta prácticamente circular ubicado a unos 1010 metros sobre el nivel del mar y por una muralla de mampostería que seguía el perímetro del cerro central. A excepción de esta última construcción, que fue erigida en periodo argárico y quedó en desuso al final de este periodo tras un incendio, el resto de zonas del poblado fueron habitadas en ambos momentos cronológicos (SÁEZ PÉREZ y ARRIBAS PALAU 1979:48).
Durante los trabajos de excavación el yacimiento fue dividido en siete zonas (Fig. 2), cinco encuadradas en el área residencial (A, B, C, D y E) y dos en la amurallada (F y G) (CONTRERAS CORTÉS 1986:241-276; MOLINA GONZÁLEZ y DORADO ALEJOS 2018:61-63). La primera campaña de excavación se desarrolló entre agosto y septiembre de 1971 bajo la supervisión de F. Molina y E. Pareja, profesores del Departamento de Prehistoria y Arqueología de la Universidad de Granada con el propósito de obtener la secuencia estratigráfica de este sitio arqueológico (MOLINA GONZÁLEZ y PAREJA LÓPEZ 1975:13). Un año después se desarrollaron dos campañas más, una durante el verano y otra entre los meses de noviembre y diciembre, con las que se puso fin a los trabajos de excavación hasta el momento (SÁEZ PÉREZ y ARRIBAS PALAU 1979: 4-6).

Fig. 2. Vista de algunos de los cortes en los que el yacimiento de la Cuesta del Negro (Purullena, Granada) fue dividido durante su excavación a comienzos de la década de los 70 del siglo XX (MOLINA GONZÁLEZ y DORADO ALEJO 2018: 62).
Desde las primeras publicaciones sobre este yacimiento, iniciadas poco después de su intervención arqueológica, las investigaciones y publicaciones no han dejado de crecer. A pesar de ello, a día de hoy tan sólo existe una obra centrada en el estudio de la fauna de este lugar arqueológico. Nos referimos al trabajo de H. D. Lauk (1976), un estudio taxonómico de una considerable cantidad de restos arqueozoológicos pertenecientes a la segunda campaña de excavación. De este modo, otro de nuestros objetivos es el de aportar mayor información al conocimiento faunístico de este yacimiento al presentar como novedad el análisis tafonómico de los restos óseos. Al mismo tiempo hemos comparado nuestros resultados con los aportados por H. D. Lauk para observar semejanzas y diferencias.
METODOLOGÍA
La metodología empleada en la identificación anatómica, taxonómica y tafonómica, así como los criterios para la determinación de edad (Tab. 1) y sexo en las especies animales determinadas es la común en este tipo de trabajos y que por repetida en múltiples publicaciones omitimos aquí (KUBASIEWICZ 1956; SCHMID y GARRAUX 1972; DRIESCH 1976; PALES y LAMBERT 1981; GRAYSON 1984; FORMING 1998; RIQUELME 1998; CHAIX y MÉNIEL 2005; WILKENS 2012; LÓPEZ PLANA 2008; ROSELL 2011; ABRIL LÓPEZ 2012; YRAVEDRA SAINZ DE LOS TERREROS 2013).
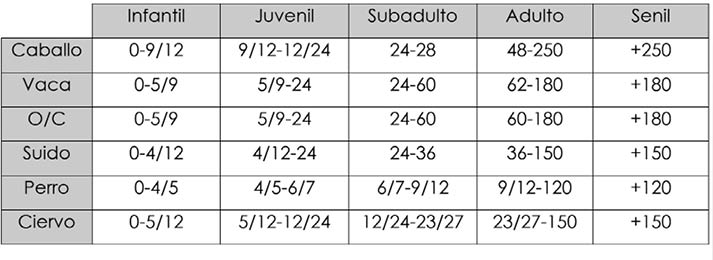
Tab. 1. Descripción de los meses de vida equivalentes a cada grupo de edad por cada mamífero identificado en la Cuesta del Negro (Purullena, Granada) (RIQUELME 1998:28).
RESULTADOS
Los materiales óseos seleccionados proceden de la campaña de excavación del verano de 1972, en concreto, de las zonas A, C, E y G. Cronológicamente se encuadran en la Fase II del periodo argárico y en el periodo de Cogotas I. Del primero de los periodos tan sólo hemos estudiado materiales del área de poblado, mientras que del segundo hemos comparado los restos de ambas zonas. De cualquier forma, en total suman 706 restos, de los cuales 442 han sido determinados (62,61%), habiendo quedado sin determinar 264 por el elevado grado de fragmentación de los materiales analizados y la falta de zonas diagnósticas (Tab. 2).
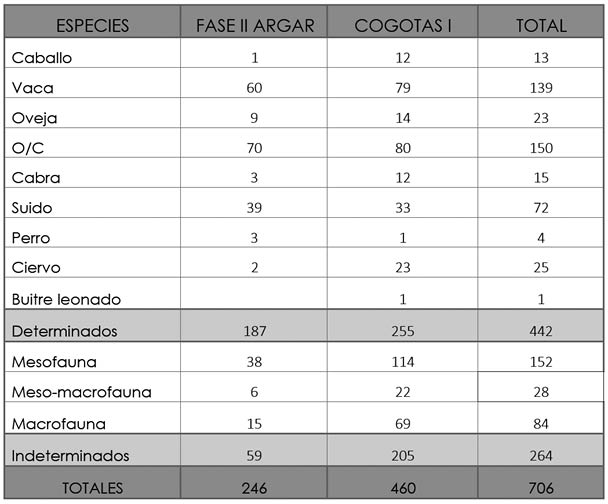
Tab. 2. Cuesta del Negro. Número de Restos contabilizados de cada taxón en los dos periodos de ocupación del yacimiento.
Respecto a las alteraciones tafonómicas, hemos contabilizado 997 ejemplos, superando al total de restos porque aunque hay algunos huesos que no evidencian marcas otros muestran más de un tipo diferente. Parece ser que las más numerosas son las vermiculaciones, es decir, alteraciones producidas por la secreción de las raíces, con 403 casos, a las que siguen las marcas provocadas por carnívoros con 134 (Tab. 3).
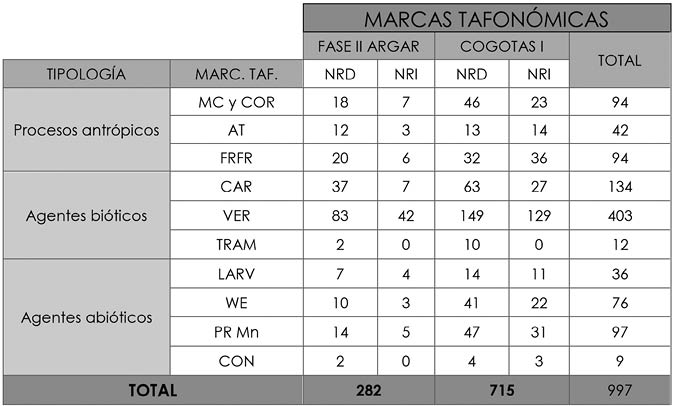
Tab. 3. Cuesta del Negro. NRD y NRI que evidencian alteraciones tafonómicas por fases cronológicas, junto a una suma total de cada tipo de marca: Marcas de corte (MC) y cortado (COR), alteración térmica (AT) y fracturas en fresco (FRFR) entre las de tipo antrópico, acción de carnívoros (CAR), vermiculaciones (VER), trampling (TRAM) y larvas (LARV) dentro de los agentes bióticos y weathering (WE), precipitaciones de manganeso (PR Mn) y concreciones (CON) como representantes de los agentes abióticos.
A continuación, desarrollaremos los resultados por periodos, primero los datos taxonómicos y tafonómicos de la Fase II del periodo argárico y posteriormente expondremos los del periodo Cogotas I.
Taxonomía y tafonomía de la Fase II del periodo argárico
De los 246 restos analizados para esta fase 187 han sido determinados taxonómicamente (76,02%), mientras que los 59 restantes (23,98%) han sido clasificados como indeterminados por las causas previamente expuestas (Tab. 4 y Fig. 3). Entre todas las especies determinadas hemos identificado tanto animales domésticos como salvajes. Dentro de los primeros encontramos caballo (Equus caballus), vaca (Bos taurus), cabra (Capra hircus) y oveja (Ovis aries) que agrupamos en ovicaprinos (O/C), cerdo (Sus domesticus) y perro (Canis familiaris). En el caso de las especies salvajes tan sólo hemos diferenciado ciervo (Cervus elaphus), aunque seguramente parte de los restos de suidos pertenezcan a jabalí (Sus scrofa). Cabe destacar que tampoco hemos diferenciado entre équidos salvajes y domésticos (Tab. 5).
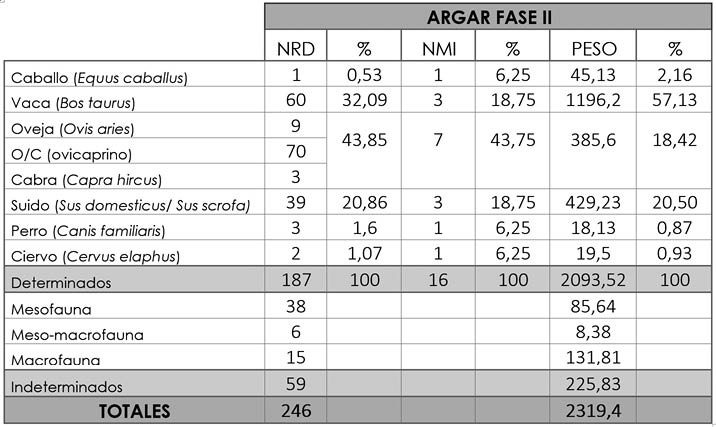
Tab. 4. Cuesta del Negro. Fase II argárica. NR, NMI y peso, junto a sus porcentajes, de los taxones determinados y de los tres grupos de indeterminados.
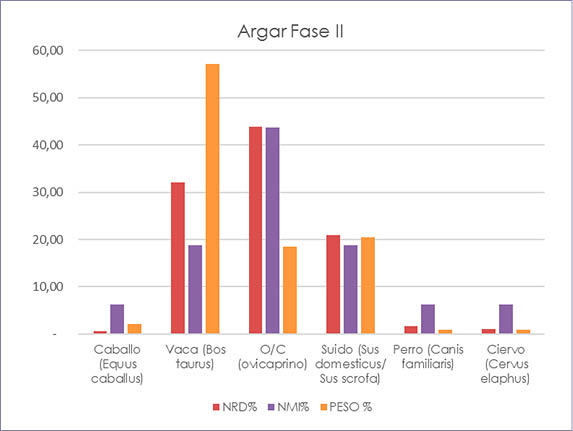
Fig. 3. Cuesta del Negro. Fase II argárica. Gráfico con los porcentajes del NRD, NMI y peso de los animales determinados.
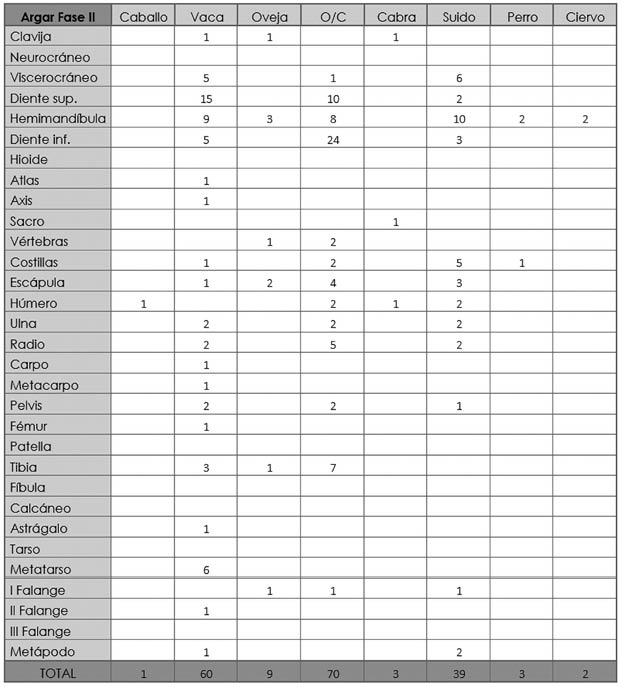
Tab. 5. Cuesta del Negro. Fase II argárica. Desglose anatómico de las especies animales determinadas.
Entre todas estas especies los huesos más numerosos son los de ovicaprinos con 82 restos (43,85%), 9 de los cuales pertenecen a oveja y 3 a cabra. Estos restos óseos provienen de un número mínimo de 7 individuos (43,75%) y, en cuanto al peso, están en tercer lugar con 385,6 g. (Tab. 4). La parte anatómica más representada es la craneal (58,54%) por la gran cantidad de dientes aislados contabilizados, a la que le sigue la apendicular (34,15%) y la axial (7,32%). Hemos documentado individuos infantiles, juveniles, subadultos y adultos, siendo estos últimos los más copiosos.
El segundo taxón más numeroso con respecto al NRD es la vaca con 60 restos (32,09%), que suponemos pertenecientes a un mínimo de 3 animales (18,75%). Esta especie constituye el mayor aporte cárnico con 1196,2 g. (57,13%) (Tab. 4). De nuevo y por el mismo motivo, la parte esquelética con mayor representatividad es la craneal (58,33%), seguida de la apendicular (36,67%) y la axial (5%). Igual que en los ovicaprinos, la edad adulta es la más numerosa, aunque también encontramos individuos juveniles y subadultos.
El tercer grupo más abundante en número de restos son los suidos con 39 huesos (20,86%) provenientes mínimo de 3 individuos (18,75%), si bien a nivel de biomasa ocupan el segundo lugar con 429,23 g. (20,5%) (Tab. 4). Como en los dos casos previos, la parte craneal es la más habitual (53,85%), seguida por la apendicular (33,33%) y por la axial (12,82%). A diferencia de los taxones anteriores, los ejemplares juveniles y subadultos son los más abundantes (Fig. 4).

Fig. 4. Radio de un suido subadulto de la Fase II del periodo argárico alterado por larvas en la mayor parte de su cortical. Elaboración propia.
En esta fase tan sólo hemos encontrado un hueso de caballo (0,53% del NRD y 6,25% del NMI) con un peso de 45,13 g. (2,16%) (Tab. 4). Este resto ha sido determinado como una epífisis proximal de un húmero izquierdo de un individuo adulto.
En el caso del perro hemos contabilizado tres restos (1,6%), dos mandíbulas y un fragmento de costilla procedentes de un número mínimo de un individuo, posiblemente de edad adulta. Estos huesos tienen un peso de 18,13 g. (0,87%) (Tab. 4).
Por último, de ciervo hemos registrado dos restos (1,07%), dos ramas mandibulares de un individuo mínimo (6,25%) y con un peso de 19,5 g. (0,93%) (Tab. 4).
En cuanto a la tafonomía, del conjunto de huesos de esta fase cronológica únicamente el 59,89% del NRD presenta alteraciones. El tipo de marca tafonómica más abundante es la producida por agentes bióticos con un 59,52%, siendo las más habituales las vermiculaciones (40,49%). Le siguen las marcas provocadas por carnívoros con un 18,05%, entre las que se han contabilizado 29 ejemplos de furrowing (mordisqueo) y 20 de pits (punzadas). Finalmente, entre los agentes biológicos tenemos las marcas producidas por larvas (Fig. 4) que representan un porcentaje de 3,41% y el trampling (pisoteo) con un 0,98% (Tab. 6).
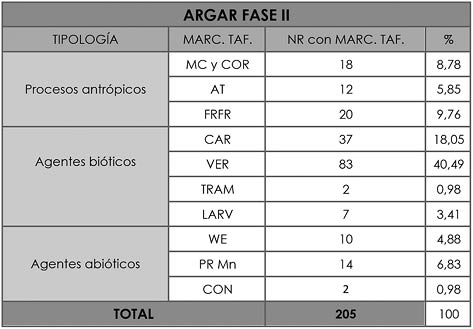
Tab. 6. Cuesta del Negro. Fase II argárica. Tipos de alteraciones tafonómicas y el número de veces que se han identificado en el NRD; así como el porcentaje que cada una de estas marcas supone para el total de alteraciones de esta fase (205).
El siguiente de los grupos en abundancia es el de marcas antrópicas con un 24,39% (Tab. 6). Las fracturas en fresco suponen el 40% de este tipo de procesos tafonómicos, las marcas de corte y cortado un 36% y la alteración térmica un 24%. La ausencia de huesos calcinados, la coloración gradual o diferencial y la contabilización de 4 casos de cocción (Fig. 5) hace plantear que estas alteraciones fueron producidas durante el cocinado, cuando el hueso aún tenía carne, y no a causa de los procesos de destrucción de residuos (CÓRDOBA SÁNCHEZ 2017:11).

Fig. 5. Detalle de la epífisis proximal de un metatarso de vaca cocido perteneciente a la Fase II argárica. Elaboración propia.
En último lugar encontramos las alteraciones causadas por agentes abióticos, y entre ellas sobresalen los huesos con precipitaciones de manganeso (6,83%), a continuación encontramos los restos con weathering o exposición subaérea (4,88%) y, finalmente, las concreciones (0,98%) (Tab. 6).
Taxonomía y tafonomía de la fase Cogotas I
Los 460 restos analizados para este periodo proceden tanto del área inferior (362 huesos) como de la superior (93). De todos ellos únicamente han podido ser determinados 255 (55,43%), quedando sin identificar 205 fragmentos óseos (44,57%) (Tab. 7 y Fig. 6). En esta fase cronológica volvemos a encontrar los mismos taxones que en el periodo previo, a excepción de una primera falange de buitre leonado (Gyps fulvus) de edad adulta. Este hueso supone un 0,39% del NRD y su peso es de 1,66 g. (0,04%) (Tab. 7 y Tab. 8).
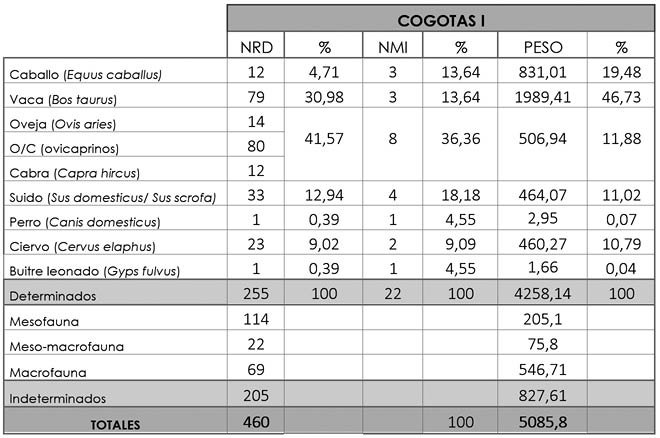
Tab. 7. Cuesta del Negro. Fase Cogotas I. NRD, NMI y peso, junto a sus porcentajes, de los taxones determinados y de los tres grupos de fauna indeterminada.
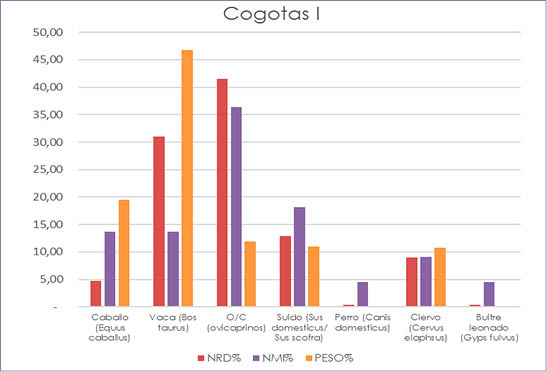
Fig. 6. Cuesta del Negro. Fase Cogotas I. Gráfico a partir de los porcentajes del NRD, NMI y peso de las especies determinadas
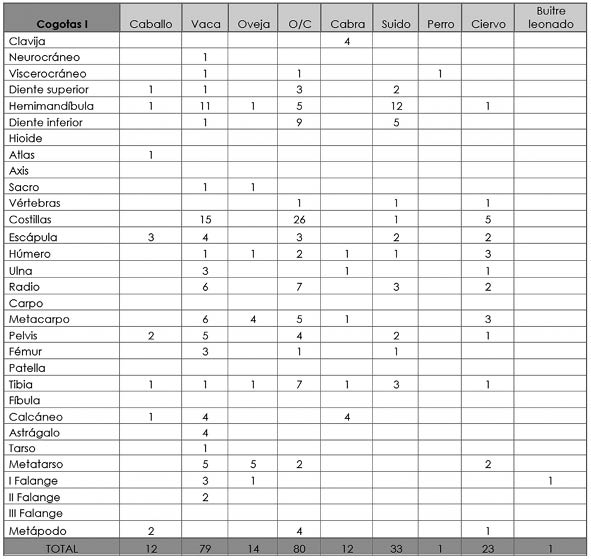
Tab. 8. Cuesta del Negro. Fase Cogotas I. Desglose anatómico de las especies animales determinadas.
Como ocurría en el periodo anterior, el NRD lo encabezan los ovicaprinos con 106 restos (41,57%), de los que 14 se identificaron como oveja y 12 como cabra. El NMI para este grupo es de 8 (36,36%) y su peso de 506,94 g. (11,88%) lo coloca en tercer lugar tras la vaca y el caballo, superando en esta ocasión a los suidos a nivel de biomasa (Tab. 7). El 51,89% de los huesos son apendiculares, el 26,42% axiales y el 21,7% craneales. Como ocurría en el periodo argárico, la mayor parte de los ejemplares llegaron a edad adulta, si bien también se han reconocido individuos infantiles, juveniles y subadultos.
Continuando con la misma dinámica, las vacas se sitúan en el segundo puesto en NRD con 79 huesos (30,98%) procedentes de un número mínimo de 3 (13,64%). Como ya hemos adelantado, es el taxón que mayor cantidad de carne proporcionó a los habitantes de la Cuesta del Negro con 1993,62 g. (46,73%) (Tab. 7). Al igual que los ovicaprinos, la parte anatómica más numerosa es la apendicular (60,76%), tras ella la axial (20,25%) y finalmente la craneal (18,99%). La edad de sacrificio sigue el mismo patrón que el de esta especie previa, con una predilección por animales adultos (Fig. 8), consumiéndose de igual modo individuos juveniles y subadultos.
El tercer grupo más numeroso, de nuevo, es el de los suidos con un total de 33 fragmentos óseos (12,94%) posiblemente obtenidos de un mínimo de 4 individuos (18,18%). En cuanto al peso, está en cuarto lugar con 464,07 g. (11,02%), muy cercano al ciervo (460,27 g., un 10,79%) (Tab. 7) que presentaremos a continuación. Los huesos más cuantiosos son los craneales (57,58%), seguido de los apendiculares (36,36%) y axiales (6,06%). En contraste con el periodo anterior, la edad de sacrificio más común es la adulta, a pesar de que los ejemplares infantiles y subadultos se siguen consumiendo.
La presencia del caballo se intensifica considerablemente en relación a la fase anterior, con 12 huesos (4,71%) de un mínimo de individuos de 3 (13,64%). A pesar de su escasez en relación al NRD, ocupa el segundo puesto en biomasa con 831,01 g. (19,48%) (Tab. 7). Los huesos de caballo más numerosos son los apendiculares (75%), después los craneales (16,67%) y, por último, los axiales (8,33%). Excluyendo un hueso de un individuo subadulto, los demás procedían de animales adultos (Fig. 7).

Fig. 7. Calcáneo de caballo con abundantes marcas de corte producto del proceso de desarticulación y furrowing en su parte superior. Procedente del área inferior del poblado. Elaboración propia.
En esta fase tan sólo hemos contabilizado un hueso de perro (0,39%), un arco cigomático derecho de un animal adulto con un peso de 2,59 g. (0,07%) (Tab. 7).
De gran interés es el incremento del ciervo con respecto al periodo argárico, con una suma total de 23 fragmentos (9,02%) de un número mínimo de dos (9,09%). Su peso es de 460,27 g. (10,79%) (Tab. 7) y nuevamente, la mayoría de los huesos son apendiculares (69,57%), le siguen los axiales (26,09%) y, en última instancia, los craneales (4,35%), una única rama mandibular de un ejemplar juvenil. A excepción de este hueso, los restantes pertenecían a individuos adultos.
Comparando las dos áreas del poblado se aprecia que los ovicaprinos representa un porcentaje mayor para el área superior (54,9%) que para la inferior (38,24%), como también ocurre con los suidos (15,69% y 12,25% respectivamente). Para el resto de taxones se produce el fenómeno contrario (Tab. 9). Estas diferencias en volumen de material, o bien de presencia de especies entre las dos partes del poblado, son bastante habituales entre los asentamientos argáricos (LULL et al. 2010:18).
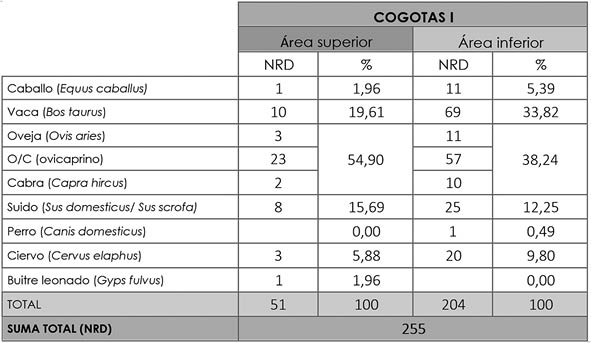
Tab. 9. Cuesta del Negro. Fase Cogotas I. NRD de cada especie determinada y su porcentaje correspondiente separando el área superior e inferior del yacimiento.
A nivel tafonómico parece ser que el porcentaje de restos alterados asciende al ser comparado con la fase argárica, con un 85,10%; siendo superior en el área de poblado que en la fortificada: 86,27% y 80,39% en ese orden.
Las alteraciones más numerosas son las de tipo biótico con 277 casos, y entre estas las vermiculaciones una vez más con un 30% en el área superior y 37,08% en la inferior. Las segundas las forman las marcas producidas por carnívoros, que significan un 16,67% de las alteraciones del área superior y un 14,59% de las de la inferior. Entre ambas zonas se han contabilizado 62 marcas defurrowing (Fig. 7) y 32 pits.En el caso del trampling existe una clara diferenciaentre el área superior (7,78%) y la inferior (0,91%). Por último, la cantidad de marcas de larvas es semejante en ambos puntos: 3,33% en la zona superior y 3,34% en la inferior (Tab. 10).
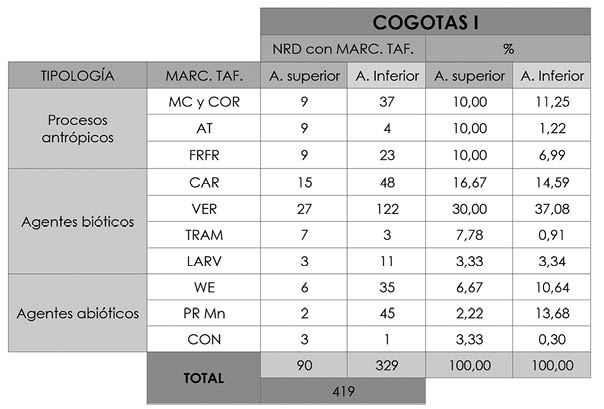
Tab. 10. Cuesta del Negro. Fase Cogotas I. Tipos de alteraciones tafonómicas y la cantidad de huesos con las mismas dentro del NRD de ambas áreas del poblado; así como el porcentaje que cada una de estas marcas supone en función del total de marcas de cada zona (90 de la superior y 329 de la inferior).
El segundo grupo en abundancia lo componen las marcas antrópicas y, entre ellas, las marcas de corte y cortado (Fig. 7) con valores muy similares en ambas áreas, con 10% en la defensiva y 11,25% en la de hábitat. Existen algunas diferencias en la frecuencia de las fracturas en fresco, siendo mayor en la zona elevada (10%) que en la inferior (6,99%). De igual modo ocurre con las alteraciones térmicas al mostrar un 10% en esa primera área y tan solo un 1,22% en la segunda (Tab. 10). Al igual que en la fase argárica, parece que estas alteraciones por fuego tienen un origen culinario; si bien en esta ocasión hemos hallado 3 huesos calcinados.
En último lugar se colocan las alteraciones abióticas. Es notable el elevado porcentaje de precipitaciones de manganeso (Fig. 8) entre todos los huesos alterados del área inferior (13,68%) si son comparados con los de la superior (2,22%). De forma similar, en la zona de hábitat observamos que los porcentajes de weathering son mayores (10,64%) que en la fortificada (6,67%). Finalmente, contamos con tan sólo un 3,33% de concreciones en el área superior y un 0,3% en la inferior (Tab. 10).

Fig. 8. Vista lateral de un astrágalo de un bóvido adulto alterado por precipitaciones de manganeso. Procedente del área inferior del poblado. Elaboración propia.
CONCLUSIONES
Como hemos podido apreciar, en ambos periodos cronológicos son los animales domésticos los más numerosos, ocupando el primer puesto en abundancia los ovicaprinos. La presencia de prácticamente todas las partes anatómicas de este grupo taxonómico y la evidencia de alteraciones antrópicas en los mismos nos indica un aprovechamiento cárnico al máximo de estos animales. La misma información puede extraerse de la preponderancia en ambos periodos de los ejemplares adultos. Mientras que los individuos de menor edad fueron aprovechados en exclusiva por su valor cárnico los adultos pudieron ser explotados para la obtención de productos secundarios como lana en el caso de las ovejas o de leche en las hembras, así como el mantenimiento del relevo generacional y, finalmente, para su carne. A pesar de que el grado de fragmentación de la muestra nos ha impedido precisar el sexo, H. D. Lauk (1976) sí logró hacer esta identificación, mostrando una preponderancia de las hembras (LAUK 1976:41).
La segunda especie más numerosa en el NRD y la primera en aporte de biomasa es la vaca en ambos periodos. Como en el grupo anterior, se observa un aprovechamiento intenso de todas sus partes anatómicas que evidencian marcas de tipo antrópico. La preferencia de individuos adultos muestra el interés por obtener un mayor rendimiento cárnico de estos individuos, pero también de su leche y de su fuerza como animal de tiro o carga. Entre los restos óseos analizados no hemos reconocido patologías óseas que respalden esta idea, pero H. D. Lauk sí pudo identificar este tipo de malformaciones en un metatarso. Asimismo, observó que en ambas fases las hembras duplicaban a los machos (LAUK 1976:30-32).
Hemos podido observar, al igual que lo hizo H. D. Lauk, que la presencia de caballo fue considerablemente mayor durante el periodo de Cogotas I (LAUK 1976:103). Llegó a suponer el segundo taxón en peso y, aunque pudo haberse utilizado por su fuerza como animal de tiro, de carga o, incluso, como elemento de poder (LUCAS PELLICER y RUBIO DE MIGUEL 1986-1987:438), su valor bromatológico es innegable (Fig. 7). En todo caso, para cualquiera de estos fines era necesario mantener al animal con vida el máximo tiempo posible, y por ello prácticamente todos los individuos estudiados fueron sacrificados en edad adulta.
Poco podemos decir del uso del perro en este asentamiento por la escasez de materiales óseos identificados y la ausencia de marcas antrópicas. No obstante, podemos deducir su presencia a partir de las marcas de carnívoros que consideramos mayoritariamente provocadas por esta especie al tener mayor facilidad de acceso al interior del poblado (CÓRDOBA SÁNCHEZ 2017:12). Como vimos, este tipo de alteración es la segunda más abundante tras las vermiculaciones. La marcada actividad de las raíces se debe a que los estratos de los que proceden los restos son muy superficiales.
Los suidos son el tercer grupo más numeroso en NRD en ambas fases y, si bien durante el periodo de Cogotas I a nivel de biomasa retroceden frente al caballo su papel dentro de la dieta de los habitantes de este poblado es indudable, como demuestran las alteraciones antrópicas y la representación de la generalidad de sus partes anatómicas. Así, los suidos domésticos serían criados con este fin, apreciándose una primera preferencia por individuos juveniles y subadultos durante la fase argárica y por los adultos durante el periodo Cogotas I. Es probable que este cambio en la edad de sacrificio nos esté mostrando un aumento de la caza del jabalí, al preferir la captura de los ejemplares más voluminosos, que serían por lo general adultos. Al mismo tiempo, en esta última etapa crece el número de ciervos en el poblado, principalmente adultos y machos (LAUK 1976:67-68). A partir de esta información podemos confirmar lo que ya se ha advertido en otros asentamientos del Bronce Final, el crecimiento de la actividad cinegética en este periodo final.
Curiosamente, la especie más recurrentemente capturada en la Cuesta del Negro fue el conejo (Oryctolagus cuniculus) aunque en nuestra muestra no se haya documentado ni un solo hueso de este taxón. De igual modo, H. D. Lauk identificó muchas más especies que no están presentes entre los restos que hemos analizado: cabra montés, liebre, corzo, lince, zorro, lobo, cigüeña blanca, buitre egipcio, búho real, etc. (LAUK 1976:103).
No queremos finalizar este artículo sin destacar algunas diferencias tafonómicas entre el área de hábitat y la fortificada y entre los dos periodos de ocupación. Si observamos el porcentaje de restos alterados por weathering y con precipitaciones de manganeso en la fase argárica seleccionada, cuyos materiales proceden del área residencial, apreciamos que los huesos fueron enterrados de forma rápida y que los niveles de humedad no eran excesivamente altos. Al comparar estos valores con el área inferior del periodo posterior observamos que la presencia de estas marcas crece porcentualmente hablando. Además, estos niveles son igualmente mayores en el área inferior que en la superior del periodo Cogotas I, probablemente por la exposición de los restos a las inclemencias del tiempo durante más tiempo y porque el grado de humedad era superior.
También apreciamos contrastes en las alteraciones térmicas, más habituales en el área fortificada, al igual que los restos óseos con marcas de trampling, seguramente por un mayor tránsito de personas y animales por este lugar.
En definitiva, la reducida muestra analizada hace de este trabajo un acercamiento a la realidad faunística de la Cuesta del Negro, información que hemos sumado a la ofrecida por H. D. Lauk en 1976. Hemos observado que la cabaña ganadera jugó un papel primordial en la economía de estas personas. Para algunos autores como F. Molina González y E. Pareja López (1975:55-56) o F. Contreras Cortés (1986:285) la trashumancia, la búsqueda de tierras favorables para el desarrollo de la actividad ganadera, sería un factor esencial en la llegada de las poblaciones meseteñas de Cogotas I a la Cuesta del Negro. Sin embargo, otros investigadores consideran que no existen suficientes datos para correlacionar una actividad económica determinada como la trashumancia con la cercanía de los yacimientos a las vías de tránsito, tan sólo podemos confirmar la existencia de contactos entre distintos grupos poblacionales (BLANCO GONZÁLEZ y ESPARZA ARROYO 2019:78). En el caso concreto de la Cuesta del Negro se ha estimado innecesario la movilización del ganado a una distancia tan lejana (ABARQUERO MORAS 2005:423-437) al contar con pastos suficientes en los terrenos ubicados antes de llegar a este enclave (BLANCO GONZÁLEZ y ESPARZA ARROYO 2019:70). Independientemente de ello, la ganadería fue un elemento clave en la economía de la Cuesta del Negro, asentamiento que sigue siendo un referente en el estudio del tránsito del Bronce Pleno al Bronce Tardío en el sureste peninsular.
BIBLIOGRAFÍA
ABARQUERO MORAS, F. J. (2005): Cogotas I. La difusión de un tipo cerámico durante la Edad del Bronce, Junta de Castilla y León, Valladolid, 2005.
ABRIL LÓPEZ, D. (2012): Contextosarqueológicos de la actividadmetalúrgicaen el suroeste de la PenínsulaIbérica (III Milenio A.N.E.): la aplicación de análisiszooarqueológicosmultivariantes, espaciales y cuantitativos para la explicación de las relacionessociales, Tesis Doctoral, Universidad de Huelva, Departamento de Historia I, Huelva, 2012, pp. 37-97.
ARANDA JIMÉNEZ, G. (2015): Resistencia e involución social en las comunidades de la EdaddelBronce del sureste de la PenínsulaIbérica, Trabajos de Prehistoria72, 1, 2015, pp. 126-444.
BLANCO GONZÁLEZ, A. y ESPARZA ARROYO, Á. (2019): Conectividad en la Edad del Bronce del occidente de la peninsula ibérica. Examinando la relación entre sitios y vías pecuarias mediante SIG, Trabajos de Prehistoria 76, 1, 2019, pp. 67-83.
CASTRO, P. V., CHAPMAN, R. W., GILI, S., LULL, V. y MICO, R. (1999): Agricultural production and social change in the Bronze Age of southeast Spain: the Gatas Project, ANTIQUITY 282, Oxford, 1999, pp. 846-856.
CHAIX, L. y MÉNIEL, P. (2005): Manual de Arqueozoología, Ariel Prehistoria, Barcelona, 2005.
CONTRERAS CORTÉS, F. (1986): Aplicación de métodosestadísticos y analíticos a los complejos cerámicos de la Cuesta del Negro (Purullena, Granada). Tesis Doctoral, Universidad de Granada, Granada, 1986, pp. 233-288.
CÓRDOBA SÁNCHEZ, A. (2017): Identificación taxonómica y análisis tafonómico de restos faunístico de la Motilla del Azuer (Daimiel, Ciudad Real), Arqueología y Territorio 14, 2017, pp. 1-18.
DRIESCH, A. von den (1976): A Guide to the Meoasurement of Animal Bones from Archaeological Sites, Peabody Museum Bulletin I, Harvard, 1976.
FORMIGN, C. (1998): Atlas osteológico digital: diferencias entre cabras, ovejas y rebecos, Universidad Autònoma de Barcelona, 1998.
GRAYSON, D.K. (1984): Quantitative Zooarchaeology. Topics in the Analysis of Archaeological Faunas, Academic Press Inc. Orlando, Florida, 1984, pp. 172-174.
JOVER MAESTRE, F. J., LORRIO ALVARADO, A. y DÍAZ TENA, M. de los A. (2016): El Bronce Final en el Levante de la península ibérica: bases arqueológicas y periodización, Complutum 27, 1, 2016, pp. 81-108.
KUBASIEWICZ, M. (1956): O metodyce badán wykopaliskowichszcaztkówkostnychzwierzecych, MaterialyZachodnio-Pomorskie 2, 1956, pp. 235-244.
LAUK, H.D. (1976):Tierknochenfunde aus bronzezeitlichen Siedlungen bei Monachil und Purullena (Prov. Granada), Studien über frühe Tierknochenfunde von der Iberischen Halbinsel 6, 1976, pp. 21-47.
LÓPEZ PLANA, C. y MAYOR APARICIO, P. (Dir.) (2008): Atlas de Osteología de los Mamíferos Domésticos, Unitatd’Anatomia i Embriologia, Departament de Sanitat i d’AnatomiaAnimals, Facultat de Veterinària, UniversitatAutònoma de Barcelona, 2008. https://veterinariavirtual.uab.cat/anatomia/osteologia/Atlas_Virtual/primera.html
LUCAS PELLICER, M.R. y RUBIO DE MIGUEL, I. (1986-1987): Introducción del caballo como animal de montura en la Meseta: Problemática, Zephyrus 39-40, Ediciones Universidad de Salamanca, Salamanca, 1986-1987, pp. 437-444.
LULL, V., MICÓ PÉREZ, R., RIHUETE HERRADA, C. y RISCH, R. (2010): Las relaciones políticas y económicas de El Argar, Menga1, 2010, pp. 11-36.
MOLINA GONZÁLEZ, F. y CÁMARA SERRANO, J.A. (2009): La cultura argárica en Granada y Jaén, En los confines del Argar: Una cultura de la Edad de Bronce en Alicante (M. S. Hernández Pérez, J. A. Soler Díaz y J. A. López Padilla, Eds.), Museo Arqueológico de Alicante, Alicante, 2009, pp. 196-223.
MOLINA GONZÁLEZ, F. y DORADO ALEJOS, A. (2018): Cuesta del Negro (Purullena, Granada), Cuadernos Técnicos de Patrimonio 7: Yacimientos arqueológicos y artefactos. Las colecciones del Departamento de Prehistoria y Arqueología (I) (F. Contreras Cortés, A. Dorado Alejos, Coord.), Universidad de Granada, 2018, pp. 61-63.
MOLINA GONZÁLEZ, F. y PAREJA LÓPEZ, E. (1975): Excavaciones en la Cuesta del Negro (Purullena, Granada). Campaña de 1971, Excavaciones Arqueológicas en España 86, 1975, Madrid.
PALES, L. y LAMBERT, C. (1971-1981): Atlas ostéologiquepour servir à l’identification des mammifères du Quaternaire, Editions du Centre national de la recherchescientifique, Paris, 1971-1981.
PÉREZ IBÁÑEZ, S. (2011): Los punzones metálicos en los contextos funerarios argáricos: Los casos de Fuente Álamo (Cuevas de Almanzora, Almería) y la Cuesta del Negro (Purullena, Granada), El Futuro del Pasado 2, 2011, pp. 63-78.
RIQUELME, J.A. (1998): Contribución al estudio arqueofaunístico durante el Neolítico y la Edad del Cobre en las Cordilleras Béticas: el yacimiento arqueológico de los Castillejos en las Peñas de los Gitanos, Montefrío (Granada),Tesis Doctoral microfilmada, Universidad de Granada, Granada, 1998, pp. 23-44.
ROSELL, J. (2001): Patronsd’aprofitament de les biomassesanimalsdurant el pleistocène Inferior i Mig (Sierra de Atapuerca, Burgos) i Superior (Abric Romaní, Barcelona), Tesis Doctoral, Universitat Rovira i Virgili, Tarragona, 2001, p. 338.
SÁEZ PÉREZ, L. y ARRIBAS PALAU, A. (1979): El complejo de fortificaciones del poblado de la Edad de Bronce de la “Cuesta del negro” de Purullena (Granada), Memoria de Licenciatura, Universidad de Granada, Granada, 1979.
SCHMID, E. y GARRAUX, O. (1972): Atlas of Animal Bones for Prehistorians, Archaeologists and Quaternary Geologists, Elsevier, Amsterdam, London, N. Y., 1972.
WILKENS, B. (2012): Archeozoologia. Manuale per lo studio dei resti faunistici dell’area mediterranea, 2012.
YRAVEDRA SAINZ DE LOS TERREROS, J. (2013): Tafonomía aplicada a la Zooarqueología, Universidad Nacional de Educación a Distancia, Madrid, 2013, pp. 185-287, 299-335.
* JAE-Intro. Instituto de Historia (IH-CSIC), Gr. de investigación Estructura Social y Territorio, Arqueología del Paisaje luciatinoco96@gmail.com
