
ESTUDIO ZOOARQUEOLÓGICO Y TAFONÓMICO DE LOS RESTOS ÓSEOS DE FAUNA DE LOS NIVELES MUSTERIENSES Y SOLUTRENSES DE LA CUEVA DEL HIGUERAL-GUARDIA (MÁLAGA-CÁDIZ). INFERENCIAS EN LAS ESTRATEGIAS DE SUBSISTENCIA Y EN LA PALEOECOLOGÍA
ZOOARCHEOLOGICAL AND TAPHONOMICAL STUDY OF THE FAUNAL BONE REMAINS OF THE MOUSTERIAN AND SOLUTREAN LEVELS OF THE HIGUERAL-GUARDIA CAVE (MALAGA-CADIZ). INFERENCES IN SUBSISTENCE STRATEGIES AND PALEOECOLOGY
José Alberto DELGADO ARCOS *
Resumen
En este trabajo se presentan los resultados del estudio zooarqueológico y tafonómico del material faunístico inédito de los niveles musterienses y solutrenses del yacimiento arqueológico de la cueva del Higueral-Guardia (Málaga-Cádiz), proveniente de la intervención arqueológica del Proyecto Kuretes en el año 2011. A partir de los resultados de ese análisis, se indagará en el origen de la acumulación ósea, las especies representadas y los procesos bioestratinómicos y fosildiagenéticos que han actuado sobre el registro óseo. Con ello se pretende realizar una aproximación a las estrategias de subsistencia de los grupos humanos del Pleistoceno superior en la región y otorgar al yacimiento un lugar en el marco de los yacimientos coetáneos del sur de la Península Ibérica.
Palabras clave
Zooarqueología, Tafonomía, Pleistoceno superior, Musteriense, Solutrense
Abstract
In this work are presented the results of the zooarchaeological and taphonomic study of the unpublished faunal material of the levels assigned to the Mousterian and Solutrean of the archaeological site of the cave of the Higueral-Guardia (Málaga-Cádiz), from the archaeological intervention of the Kuretes Project in 2011. From the results of that analysis, it will investigate the origin of the bone accumulation, the species represented and the biostratinomic and fossildiagenetic processes that have acted on the bone record. With this, it is intended to make an approach to the subsistence strategies of the upper Pleistocene human groups in the region and to give the site a place in the time frame of the contemporary sites in the south of the Iberian Peninsula.
Keywords
Zooarchaeology, Taphonomy, Upper Pleistocene, Mousterian, Solutrean
INTRODUCCIÓN. EL YACIMIENTO PLEISTOCENO DE LA CUEVA DEL HIGUERAL-GUARDIA
El sur de la Península Ibérica, por sus singularidades geográficas y orográficas, constituye un enclave con excelente potencial para albergar secuencias arqueológicas que permitan conocer y comprender la evolución de los grupos humanos y su entorno ecológico durante el Pleistoceno superior. La Serranía de Ronda, donde se encuentra el yacimiento objeto de este trabajo, es uno de esos entornos fundamentales para entender la dinámica subsistencial de las comunidades humanas durante el Cuaternario. Más aún en un contexto en el que la investigación es más bien escasa.
La cueva del Higueral-Guardia se ubica entre los términos municipales de Cortes de la Frontera (Málaga) y Jerez de la Frontera (Cádiz). Se encuentra a más de 400 msnm, insertada en el cerro de las Motillas, el cual forma parte del Parque Natural de los Alcornocales en la Serranía de Ronda, adscrita al sistema de las Béticas occidentales (Fig. 1) (BAENA et al. 2012: 108).
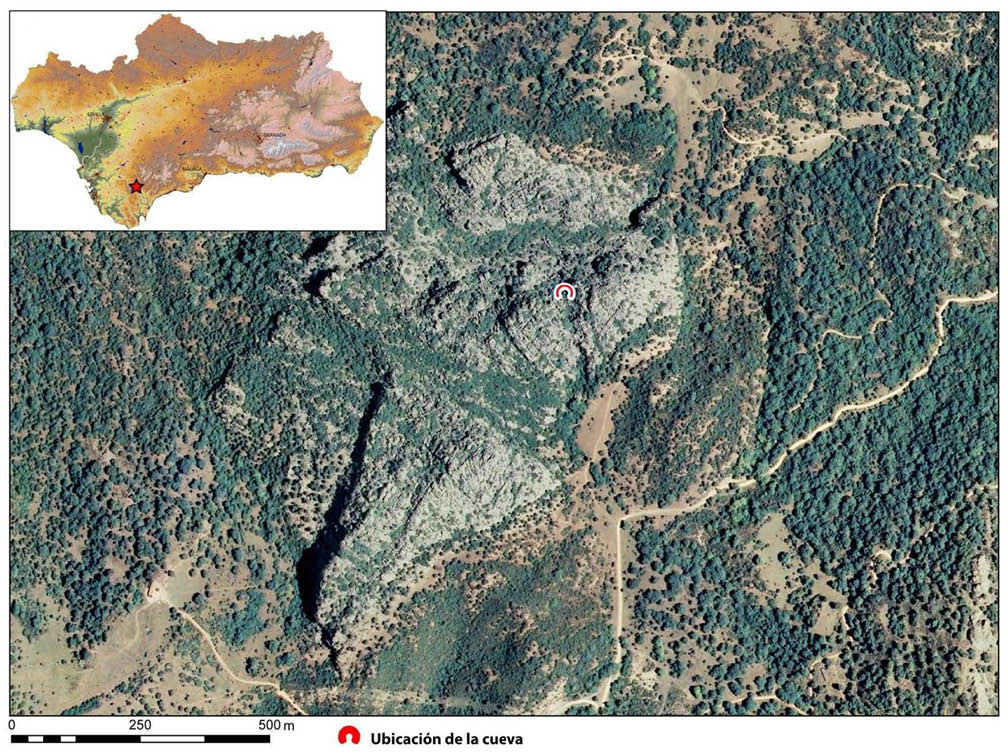
Fig. 1. Localización de la cueva del Higueral-Guardia. Extraído de BAENA et al. (2012: Figura 1).
El cerro de Las Motillas ha sido conocido desde antiguo y presenta numerosas evidencias arqueológicas, entre las que podemos destacar la conocida cueva de Las Motillas, que contiene pinturas paleolíticas atribuidas a momentos solutrenses (SANTIAGO VÍLCHEZ 1990 y 2000), la cueva del Quejigo o el abrigo del Bombín, ambas con registros arqueológicos del Paleolítico superior (GILES PACHECO et al. 1997). También se ha constatado la existencia de un asentamiento de la Prehistoria reciente en la parte alta del cerro (AGUAYO et al. 1993).
Sobre la cueva del Higueral-Guardia se han realizado dos actuaciones arqueológicas hasta el momento. La primera se trata de una reactivación de un perfil fruto del expolio por parte del Proyecto de Investigación “Prospecciones Arqueológicas Superficiales en la cuenca del río Guadalete. Análisis geocronológicos y sedimentológicos” (GILES PACHECO et al. 1997), llevándose a cabo una primera secuenciación de niveles en los que se incluía un nivel con material lítico propio de cronologías solutrenses. De dicha intervención se obtuvo también una serie de materiales faunísticos que han sido estudiados y publicados (CÁCERES y ANCONETANI 1997; CÁCERES 2003).
La segunda de esas intervenciones, y de la que salen los materiales fruto de este trabajo, es la intervención del “Proyecto Kuretes. Primeras ocupaciones humanas, evolución paleoecológica y climática del Cuaternario de las Béticas occidentales (2010-2015)” (BAENA et al., 2013), coordinado por J. Baena Preysler (Universidad Autónoma de Madrid), A. Morgado Rodríguez (Universidad de Granada) y J.A. Lozano Rodríguez (Instituto Andaluz de Ciencias de la Tierra del CSIC).
En este proyecto se realizaron tres sondeos en distintas zonas de la cueva (Fig. 2) para comprobar el potencial y el alcance de la secuencia estratigráfica, constatándose un importante expolio en los niveles superiores (BAENA et al., 2012: 111-112). Sólo en el sondeo 2, se llegó a niveles más antiguos de cronología musteriense sin alteración y es sobre el que se ha podido realizar un perfil estratigráfico (Lám. 1) extrapolable a los otros dos sondeos. El nivel 1 se corresponde a momentos recientes y a una ocupación neolítica de probable carácter funerario. Los niveles 2 y 3 albergan una serie de potentes paquetes con material adscrito al Solutrense evolucionado, con posible interestratificación de un Solutrense superior; aunque son los niveles que presentan el expolio.
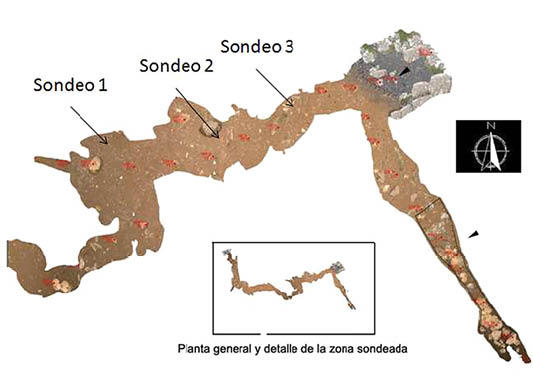
Fig. 2. Distribución de sondeos arqueológicos sobre planta. Grupo espeleológico G40. Extraído de BAENA et al. (2012: Figura 3).

Lám. 1. Perfil oeste del sondeo 2 con diferenciación de los niveles. Fuente: BAENA et al (2012: Lámina 5)..
Los niveles del 4 al 6 constituyen un hiato en el que apenas se encuentra material arqueológico. El 7 es el único del que se ha realizado una datación absoluta que arrojó una fecha de 37410±240 BP sin calibrar (Beta-318022), nos encontramos aquí en los últimos momentos del interestadial OIS 3, transición entre la glaciación Würm III y II. Por último, los niveles 8 y 9 se adscriben también a una fase musteriense pero más antigua, por la presencia en la industria del esquema operativo tipo quina, en contraste con la de tipo levallois documentada en el nivel 7 (Lám. 2) (BAENA et al. 2012:113).

Lám. 2. Núcleo levallois musteriense, procedente del nivel 7 del sondeo 2. Fuente: BAENA et al. (2012: Lámina 14).
La industria lítica obtenida el sondeo 2 ha sido estudiada y publicada (TORRES et al. 2012). El estudio de esta industria, aunque encontrada en niveles revueltos, es la que ha permitido fechar el material y los niveles de Paleolítico superior en un Solutrense superior evolucionado andaluz. Entre el repertorio se encuentran productos laminares, productos foliáceos, raspadores, núcleos laminares muy explotados, buriles y un conjunto de puntas de pedúnculo y aletas (Lám. 3) (TORRES et al. 2012:228).

Lám. 3. Esbozos y puntas finalizadas de pedúnculo y aletas halladas en los niveles solutrenses del sondeo 2. Fuente: TORRES et al. (2012: Figura 9).
MATERIALES Y METODOLOGÍA
Los materiales objeto de análisis de este trabajo son los correspondientes a la campaña de excavación del Proyecto Kuretes sobre la cueva del Higueral-Guardia en 2011. De ese material faunístico se ha hecho una selección de los niveles musterienses (7, 8 y 9 del sondeo 2) y solutrenses (2 y 3 de los tres sondeos). Esa selección se ha dividido en tres conjuntos separados: los niveles 8 y 9 musterienses, el nivel 7 musteriense y los niveles con material revuelto solutrense. Esa agrupación se basa en el hecho de que cada uno presenta evidencias para poseer entidad propia por las razones antes vistas. En total se ha estudiado un número de 1.765 restos óseos.
Se ha realizado una cuantificación de los restos recuperados a través de cálculos de abundancia relativa: el Número de Restos (NR) y el Número Mínimo de Individuos (NMI). El NR se refiere al número de restos total (YRAVEDRA 2006). Se ha aplicado tanto a determinables como indeterminables, para todas las especies y tallas y para cada elemento óseo. El NMI se ha calculado siguiendo a Brain (1969) en función del elemento óseo más numeroso, lateralidad, edad y diferencias entre sexos en el caso de que pudieran observarse.
La identificación taxonómica se ha realizado con ayuda de la colección ósea del Departamento de Paleontología de la Universidad de Granada, junto con el apoyo de los atlas osteológicos de Pales y Lambert (1971) y Schmid (1972). La avifauna ha sido determinada por el Dr. A. Sánchez-Marco, investigador del Institut Català de Paleontologia Miquel Crusafont (ICP). En los casos en los que la especie no ha podido ser identificada, se ha hecho una agrupación de los animales relacionada con su talla (RIQUELME 1998 y YRAVEDRA 2006): macromamíferos (Bos primigenius, Equus ferus), mesomamíferos (Capra pyrenaica, Sus scrofa, Cervus elaphus, Capreolus capreolus, Felis silvestris y pequeños carnívoros) y micromamíferos (lagomorfos, mustélidos, roedores). A ello se ha añadido el grupo de los macro-mesomamíferos, en el que se incluyen elementos de los que no se tiene certeza su adscripción a uno u otro grupo y las aves indeterminadas. La edad de los taxones documentados se ha establecido en función del desgaste dental (GARCÍA 2014; MOIGNE 1983) y el estado de fusión epifisaria de los huesos largos (SCHMID 1972). En cuanto a determinación sexual, apenas se han podido realizar inferencias dada la fragmentación del conjunto y la ausencia de elemento diagnósticos.
Dentro de los perfiles esqueléticos, los elementos no identificados anatómicamente se han clasificado en (BLASCO LÓPEZ 2011 y ROSELL 2001): huesos largos (estilopodios: húmero y fémur; zeugopodios: radio, ulna, tibia y fíbula; metapodios: metacarpo y metatarso; y acropodios: falanges), huesos planos (cráneo, mandíbula, vértebras, costillas, escápula, y pelvis), huesos articulares (carpales, tarsales y patelas) e indeterminados. Las secciones óseas se han clasificado en craneales (huesos del cráneo), axiales (vértebras, costillas, escápula y pelvis), apendiculares superiores (húmero, fémur, tibia y radio) y apendiculares inferiores (metapodios, falanges, carpos y tarsos) (YRAVEDRA 2006).
Por razones de tiempo y relevancia, el análisis tafonómico se ha efectuado sobre los elementos de macromamíferos, mesomamíferos y conejo. El proceso de observación de los restos óseos se ha llevado a cabo con la ayuda de lupa de mano de 40x y lupa estereoscópica, siguiendo a Blumenschine (1995).
Los patrones de fragmentación y fractura se han analizado midiendo la longitud de los restos para evaluar su intensidad y agrupándolos por medidas en centímetros (YRAVEDRA 2006). El grado de circunferencia de las diáfisis de los huesos largos se ha medido siguiendo a Bunn (1982) y establiendo cuatro grados: <25% del total de la circunferencia, 25-50%, 50-75% y más del 75%. Para estos análisis se ha hecho una división entre los huesos de macro y mesomamíferos y los huesos de conejo, para que los resultados no se vean distorsionados. Para el tipo de fractura, se han seguido los estudios de Villa y Mahieu (1991). El análisis se ha efectuado en base a la delineación con respecto al eje longitudinal hueso (longitudinal, transversal o curvado) y la superficie de fractura (suave o irregular).
Las marcas de corte se han identificado siguiendo a Binford (1981), Blasco López (2011) y Shipman y Rose (1983). Cuando ha sido posible, las marcas de corte han sido identificadas con una serie de labores antrópicas de procesado de las carcasas animales (YRAVEDRA 2006): desollado, evisceración, descuartizado, desarticulado, descarnado y limpieza del periostio. En cuanto a marcas de percusión se ha seguido para su identificación a Blumenschine y Selvaggio (1988), Fisher (1995) y Pickering y Egeland (2006). Para las alteraciones térmicas se ha hecho una gradación en función de la coloración en base a los criterios de Stiner et al. (1995). Para la distinción de las huellas de carnívoros se han analizado las superficies óseas en busca de surcos, depresiones, impresiones, perforaciones y vaciados de epífisis; siguiendo a Binford (1981), Bunn (1981) y Yravedra (2006). También se han buscado marcas de roedores (BLASCO 1992) y alteraciones por raíces o vermiculaciones (GARCIA 2014).
Respecto a alteraciones físico-químicas, se han realizado una serie de gradaciones en función de la presencia de alteración aérea o weathering (BEHRENSMEYER 1978), concreción, disolución, manganeso (GARCÍA 2014), redondeamiento (CÁCERES 1995 y 2002), pulido, trampling o pisoteo (YRAVEDRA 2006) y compactación (SHIPMAN 1981).
ANÁLISIS FAUNÍSTICO
Identificación taxonómica, edades y perfiles esqueléticos
Niveles 8 y 9 del Musteriense
Los niveles 8 y 9 musterienses han arrojado un total de 571 restos óseos (Tab. 1). De ellos se han identificado taxonómica y anatómicamente 136 (31,26% del total), ascendiendo el total de los indeterminados a 435 (79,18%). Entre el total determinados, el taxón que más restos ha aportado ha sido el conejo (Oryctolagus cuniculus), con el 89,71 % del total. Le siguen muy por debajo los ungulados representados por la cabra (Capra pyrenaica), el ciervo (Cervus elaphus) y el uro (Bos primigenius).
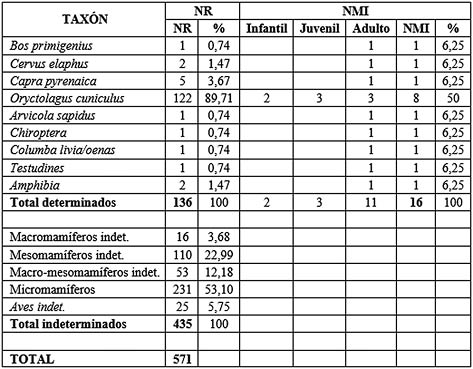
Tab. 1. Número de Restos (NR) y Número Mínimo de Individuos (NMI) de los mamíferos para los niveles 8 y 9 musterienses.
Se ha atestiguado la presencia de otros taxones de micromamíferos, reconociéndose la presencia de rata de agua (Arvicola sapidus) y una especie de quiróptero (orden Chiroptera). Las aves las representa la paloma bravía o zurita (Columba livia/oenas). Por último, se ha documentado una placa de caparazón de quelonio (Testudines) y dos elementos de anfibio (Amphibia) de los que no se ha podido especificar más.
El NMI obtenido ha sido de 16. De ellos 8 los representa el conejo (dos infantiles, dos juveniles y tres adultos) y las demás especies sólo aportan un individuo cada una.
En cuanto a los indeterminados, destaca la alta cantidad de micromamíferos (algo más del 50%), seguidos por los mesomamíferos, el grupo de macro-mesomamíferos, las aves y los macromamíferos.
Para los perfiles esqueléticos (Tab. 2) el conejo es el que arroja más información. El esqueleto apendicular inferior es el más numeroso, implicando el 50% del total de restos de conejo. Los elementos más numerosos aquí son los metápodos. El esqueleto apendicular superior lo sigue en peso, con un 28%. En esta región los huesos más numerosos son la tibia y el fémur. El craneal y el axial se sitúan al mismo nivel con valores del 11%. El único fragmento identificado de uro es una metáfisis distal de fémur. El ciervo está representado por un radio y una tibia y la cabra por varios elementos del esqueleto apendicular inferior. Los fragmentos identificados de macromamífero indeterminado son en su mayoría huesos largos sin identificar. Los elementos de mesomamífero destacan por la cantidad de elementos axiales que refleja. Esto indicaría un transporte más completo de las carcasas. Los huesos largos (esqueleto apendicular) los siguen en importancia.
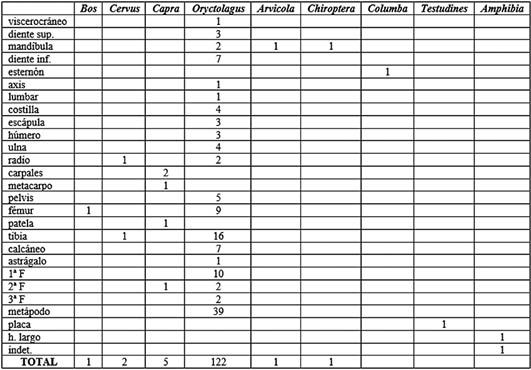
Tab. 2. Perfiles esqueléticos de los taxones documentados para los niveles 8 y 9 del Musteriense en base al NR.
Nivel 7 del Musteriense
En el nivel 7 se ha muestreado un reducido número de 194 restos (Tab. 3). De estos, ha sido posible determinar el taxón de 45 elementos (23,20 % del total), mientras que no se ha podido llegar a identificar la especie en 150 restos (77,32 %).
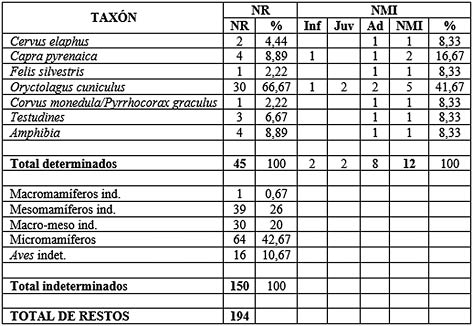
Tab. 3. Número de Restos (NR) y Número Mínimo de Individuos (NMI) (Inf: infantil, Juv: juvenil, Adult: adulto) del nivel 7 del Musteriense.
De los taxones determinados, el que más restos ha sumado para este nivel vuelve a ser el conejo con casi el 70 % del total de los determinados. Los demás taxones aparecen de manera muy reducida. Los ungulados que aparecen son la cabra y el ciervo. El único carnívoro encontrado es el gato montés (Felis silvestris). En cuanto a aves, se ha hallado un fragmento atribuible a un córvido de pequeño tamaño (Corvus monedula-Pyrrhocorax graculus). También se han encontrado restos de quelonio y anfibios a los que no se ha podido atribuir especie.
Para el NMI, de los 12 individuos contabilizados, el conejo presenta cinco de ellos, con dos adultos, dos juveniles y uno infantil. La cabra la representan un espécimen adulto y un infantil. El ciervo aporta un animal adulto. El resto de los taxones aportan un individuo cada uno.
Los grupos de talla no han compensado demasiado la exigua información aportada por los taxones. El grupo de los micromamíferos es el más numeroso con 64 restos, seguido de los mesomamíferos, los indeterminados atribuibles a macro o mesomamíferos, las aves sin determinar y un único fragmento atribuido a macromamífero.
Salvo el conejo, los demás taxones no presentan el suficiente número de restos como para realizar interpretaciones de patrones esqueléticos (Tab. 4). En el conejo la región esquelética más representada es la apendicular inferior, con un 53% del total, representada por ocho metápodos y ocho falanges. El esqueleto apendicular superior aporta un 27% al total del conjunto, incluyendo fragmentos de ulna, radio, fémur (hueso más numeroso) y tibia. El craneal y el axial aportan pocos elementos.
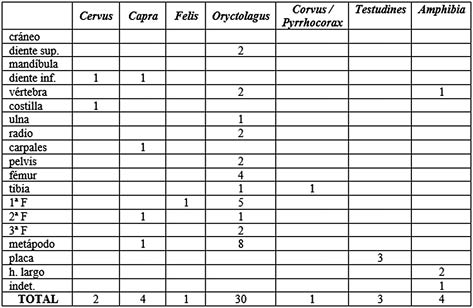
Tab. 4. Perfiles esqueléticos de los taxones documentados para el nivel 7 Musteriense en base al NR.
Niveles solutrenses
Para los niveles solutrenses, se han analizado un total de 996 restos (Tab. 5). De estos la identificación taxonómica ha sido posible realizarse sobre un total de 333 elementos (33,43 % del total), pasando a los indeterminados un número de 663 fragmentos (66,57 %). La variedad de taxones documentados en estos niveles es mayor respecto a los anteriores. Como regla general en todos los paquetes analizados, el conejo es el que más elementos óseos aporta representando un 61,86 % del total de determinados. Le siguen los ungulados, encabezados con el ciervo y la cabra montés, que aportan un 15% del total respectivamente. A nivel testimonial se ha documentado la presencia de jabalí con 3 restos. De carnívoro se ha atestiguado un resto de gato montés. Igualmente destaca la presencia de erizo.
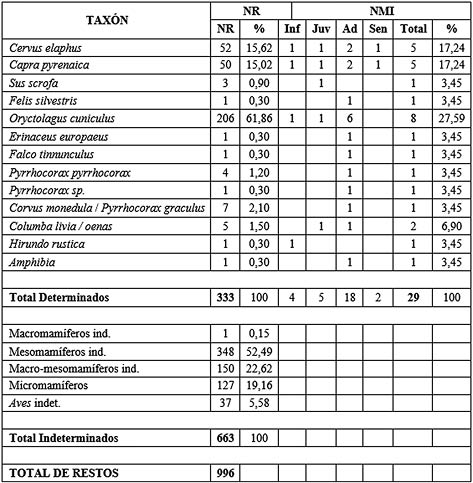
Tab. 5. Número de Restos (NR) y Número Mínimo de Individuos (NMI) (Inf: infantil, Juv: juvenil, Ad: adulto y Sen: senil) para los niveles solutrenses.
La avifauna en estos niveles aporta un número mayor de especies. Destaca el hallazgo de un fragmento de rapaz (cernícalo común, Falco tinnunculus). Más numerosos son los restos asignables a pequeño córvido (Corvus monedula-Pyrrhocorax graculus), seguido de la paloma, la chova piquirroja, un tipo de chova sin identificar y la golondrina (Hirundo rustica/daurica). Por último, se ha documentado un elemento de anfibio sin identificar.
Se han contabilizado un total de 29 individuos. La especie más numerosa la conforma el conejo con ocho ejemplares: un infantil, un juvenil y seis adultos (27,59 % del total), le siguen en importancia, el ciervo y la cabra montés, cada uno con 5 individuos: un infantil, un juvenil, dos adultos y un senil respectivamente. Para el resto de los taxones el número es de uno, excepto la paloma que aporta dos. Todos adultos menos un juvenil de jabalí y de paloma y la golondrina con un infantil.
En cuanto a los grupos de tamaño en el apartado de indeterminados, hay un destacado predominio de los mesomamíferos con 348 restos (52,49 % del total de indeterminados), que podría corresponderse con ciervo, cabra montés y jabalí. Les siguen los indeterminados atribuibles a macro o mesomamíferos con 150 restos (22,62 %). El grupo de macromamíferos está representado por un resto. Los micromamíferos representados suponen un 19,16% y la avifauna sin determinar un 5,58 %.
Los perfiles esqueléticos (Tab. 6) para estos niveles aportan más información que para los anteriores. En el ciervo las partes del esqueleto apendicular inferior son las más representadas con un 44% del total, seguidas de cerca por las craneales (42%). Menos numerosas son las pertenecientes al esqueleto apendicular superior (12 %), siendo las del esqueleto axial las menos representadas con un fragmento de pelvis. Conviene apuntar que la región craneal puede verse engrosada por la mayor presencia de dientes aislados y también de varios fragmentos de clavija. Por otra parte, la abundancia de restos procedentes de la región apendicular indica un transporte bastante completo de la carcasa animal al yacimiento.
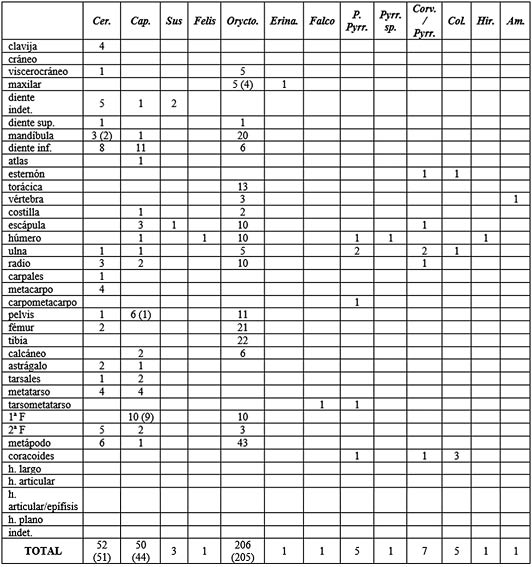
Tab. 6. Perfiles esqueléticos de los taxones documentados para los niveles solutrenses en base al NR. Entre paréntesis, variante del número de restos debido al remontaje de algunos de ellos. Cer.: Cervus; Cap.: Capra; Orycto.: Oryctolagus; Erina.: Erinaceus; P. Pyrr.: Pyrrhocorax pyrrhocorax; Pyrr. sp.: Pyrrhocorax sp.; Corv.: Corvus; Hir.: Hirundo; Col.: Columba; Am.: Amphibia.
La representación anatómica en la cabra montés muestra una predominancia, al igual que en el ciervo, del esqueleto apendicular inferior con un 48% del total. Algo más de una cuarta parte del total lo representa el esqueleto craneal, seguido por el axial que, a diferencia de lo ocurrido con el ciervo, es más numeroso con un 14% del total. El esqueleto apendicular superior queda representado con un 8% del total respectivamente. En este caso, la región craneal también se ha visto aumentada considerablemente en número por la abundancia de dientes aislados. Aunque a nivel de NR el axial es considerablemente superior al del ciervo, hay que resaltar que se ha podido remontar los restos de una pelvis de cabra montés. Los parámetros de transporte se asemejan al del ciervo con una carcasa bastante completa, destacando la región apendicular inferior.
En el conejo las partes esqueléticas quedan más repartidas, el esqueleto apendicular superior es el más numeroso con un 34% del total, seguido por el inferior con el 29% y una equitativa representación del craneal y axial entorno al 20%. Para el resto de las especies documentadas el establecimiento de conclusiones sobre su representatividad es prácticamente nulo.
Respecto a la identificación por tallas, sólo se pueden hacer inferencias en cuanto a las regiones anatómicas en los mesomamíferos por su número de restos. En este grupo, casi el 50% de los restos corresponden al esqueleto apendicular, siendo el resto completado por elementos craneales y axiales.
Estudio tafonómico
Niveles 8 y 9 del Musteriense
Patrones de fragmentación y fractura
El conjunto óseo de los niveles 8 y 9 se encuentra muy fragmentado (Fig. 3). La mayoría de los restos (67,38%) se encuentran en valores de menos de los tres centímetros de longitud. La medición del grado de circunferencia (Fig. 4) establece que el 96 % de los fragmentos diafisarios (sin contar el conejo) conservan menos del 25 % de la circunferencia. El conejo muestra valores totalmente diferentes (Fig. 4). De esta especie, el 81 % de fragmentos diafisarios conserva de manera completa la circunferencia. Estos porcentajes son normales en yacimientos con aportación antrópica de lagomorfos, puesto que su consumo es distinto. Estas aportaciones se caracterizan por una elevada fragmentación y la existencia de cilindros (en este caso de origen antrópico) de huesos largos (YRAVEDRA 2008b:86).
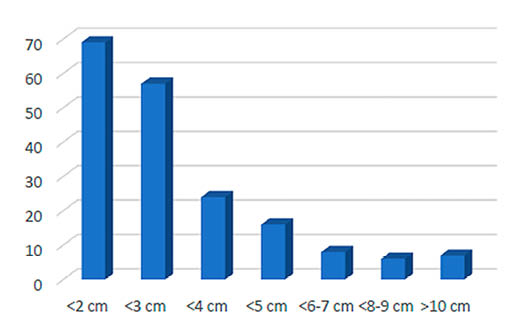
Fig. 3. Patrones de fragmentación en cuanto a longitud de los restos en base al NR para los niveles 8 y 9 musterienses, sin incluir conejo.
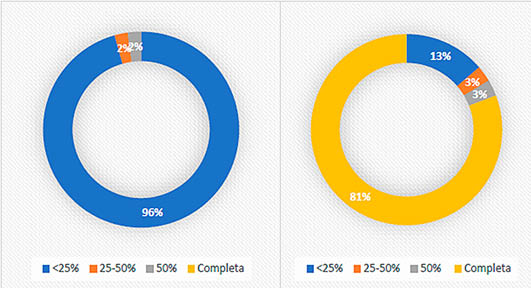
Fig. 4. Grado de circunferencia de los fragmentos diafisarios sin incluir conejo a la derecha y grado de circunferencia de los fragmentos diafisarios del conejo a la izquierda de los niveles 8 y 9 musterienses.
Los paños de fracturación (Tab. 7) nos indican que los huesos fracturados en fresco son los más numerosos con algo más del 50 % del total. La morfología de fractura (Tab. 8) determina que la más numerosa es la longitudinal con el 31,07 % del total, seguida de la oblicua con el 22,33 %. Todos los taxones muestran fracturas en fresco, siendo el que más los mesomamíferos con 25 elementos, seguida del conejo con 17. En la morfología de fractura, para la longitudinal los mesomamíferos también son los más numerosos, mientras que para la oblicua lo es el conejo.
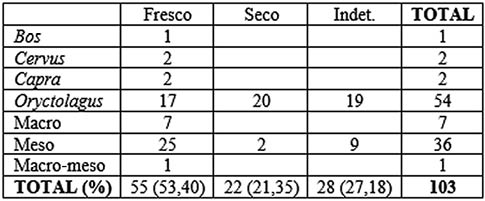
Tab. 7. Tipo de fractura sobre huesos largos en base al NR en los niveles 8 y 9 musterienses.
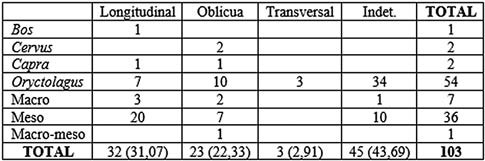
Tab. 8. Morfología de fractura sobre huesos largos en base al NR en los niveles 8 y 9 musterienses.
Alteraciones antrópicas
La actividad humana se ha visto plasmada en el conjunto de estos niveles musterienses (Tab. 9). Todos los taxones presentan evidencias de esta en mayor o menor medida. Las marcas de corte ascienden a un total de 28. La presencia de algunas de ellas en elementos del esqueleto apendicular superior e inferior indica un acceso primario a los recursos y un proceso de carnicería con un transporte bastante completo, como evidencia la presencia de elementos axiales con evidencias de corte. Las incisiones son las tipologías más abundantes en la muestra, junto con algunos aserrados, un tajo y un raspado. Muchos de los huesos presentan un número elevado de marcas, indicadores de su intenso y completo aprovechamiento. A raíz de estas marcas, se han podido distinguir una serie de labores de carnicería. La mayoría de las marcas presentes se centran en la tarea de extraer el máximo posible de carne al hueso (descarnado) (Lám. 4), habiendo algunas marcas destinadas al desollado (Lám. 5), la desarticulación (Lám. 6), la evisceración y la limpieza de periostio.
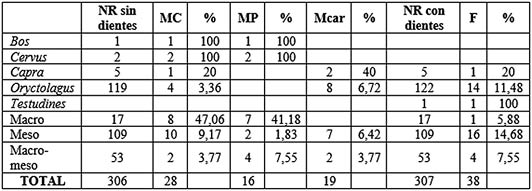
Tab. 9. Resumen de acción antrópica y de carnívoros sobre los taxones documentados en los niveles 8 y 9 del Musteriense. A efectos de porcentajes se han quitado los dientes y clavijas para las marcas de corte, de percusión y de carnívoros. MC: marcas de corte, MP: marcas de percusión, Mcar: marcas de carnívoros, F: fuego.
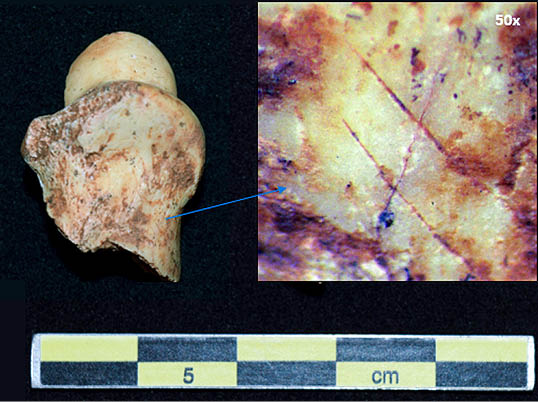
Lám. 4. Segunda falange de cabra montés con incisiones que evidencian la actividad de desollado. Fuente: elaboración propia.
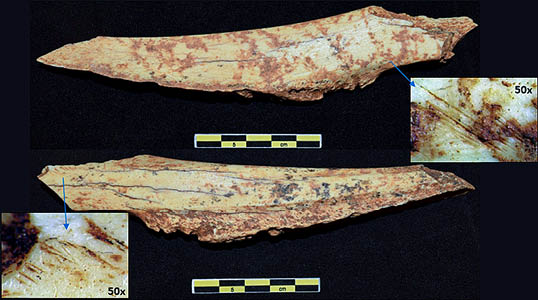
Lám. 5. Ulna de macromamífero con aserrados que muestran la actividad de desarticulado. Fuente: elaboración propia.
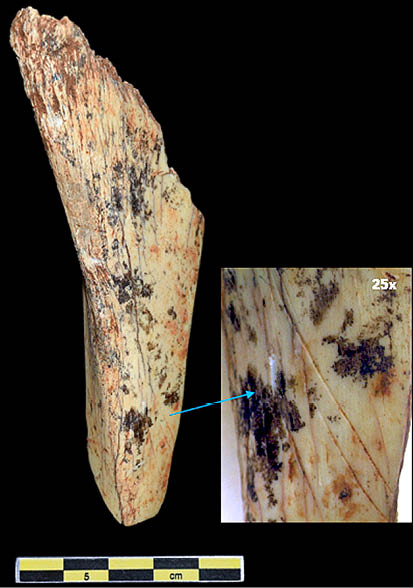
Lám. 6. Tibia de ciervo con evidencias de descarnado. Fuente: elaboración propia.
Las marcas de percusión reflejan la intencionalidad de gran parte de las fracturas buscando el acceso al tuétano. Se han contabilizado 16 evidencias de fracturación intencional, entre las que se encuentran muescas (Lám. 7), conos de percusión (Lám. 8), lascas medulares y diversos estigmas fruto de los impactos sobre la superficie ósea. Doce de los restos presentan puntos de impacto. Cuatro de ellos tienen más de uno y la tipología existente es la de puntos de impactos correlativos y consecutivos. La mayoría de ellos presentan tanto marcas de percusión como marcas de corte por toda su superficie. Este factor nos habla de patrones de consumo en los que el procesado de los recursos de las porciones óseas es máximo.
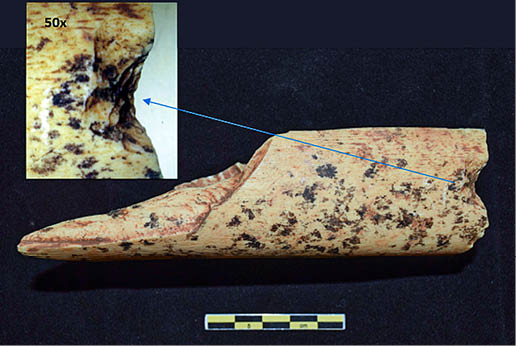
Lám. 7. Muesca sobre hueso largo de macrofauna. Fuente: elaboración propia.
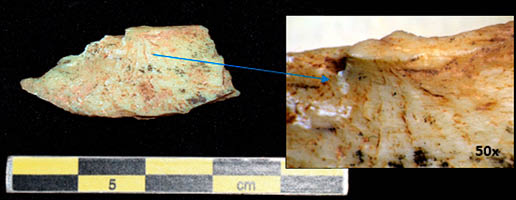
Lám. 8. Cono de percusión procedente de un elemento de macrofauna. Fuente: elaboración propia.
Los restos con evidencias de acción térmica no son demasiado numerosos. En total hay 37 (11,94 % del total de restos), siendo más destacados los que han sido quemados de manera total y con una coloración negra. Esta información contrasta con las evidencias de marcas de corte y percusión y nos hablaría de que el cocinado de la carne, en base a los restos disponibles, es escaso.
Alteraciones por otros agente biológicos
Los carnívoros (Lám. 9) han incidido en cierta medida en el registro óseo de estos niveles. Se han encontrado 19 restos con evidencias (6,21 % del total), presentando dos de ellos superposición sobre huellas antrópicas. Las improntas de carnívoros que encontramos son impresiones, surcos y depresiones. Por las dimensiones de las marcas de diente parece tratarse de pequeños carnívoros; que difícilmente pueden transportar animales de gran talla.

Lám. 9. Metacarpo de cabra montés con presencia de improntas de dientes de carnívoro. Fuente: elaboración propia.
Se han encontrado dos restos con evidencias de marcas de roedores. Ambos son diáfisis de huesos largos de animales de talla media. La acción de raíces también es muy escasa. En total se han contado 19 restos con estas evidencias de manera somera.
Alteraciones físico-químicas
Los restos óseos presentan un buen estado de conservación. La afección que más incide sobre ellos con diferencia es la del manganeso, que afecta a 146 restos (47,25 % del total). El weathering (alteración subaérea) tiene algo de peso, incidiendo sobre 91 fragmentos (29,45 %). La concreción está por debajo de esos valores, la encontramos en 74 restos (23,95 %) y apenas afecta a las superficies óseas. El redondeamiento y pulido, relacionados con la abrasión física y la acción del agua, se encuentran escasamente representados, al igual que la compactación por el sedimento. El trampling es el agente físico menos documentado, presentando sólo dos restos.
Nivel 7 del Musteriense
Patrones de fragmentación y fractura
La muestra del nivel 7 se encuentra también muy fragmentada, la gran mayoría de fragmentos (sin incluir el conejo) se encuentran por debajo de los tres centímetros de longitud. El grado de circunferencia es inferior al 25% en todos ellos. En el conejo la gran mayoría de las diáfisis conservan su circunferencia completa. Se han podido identificar pocas evidencias de fracturación. Se han documentado siete fracturas en fresco: una sobre un elemento de conejo, otra sobre un metápodo de talla grande, en un hueso largo de talla pequeña y en cuatro de talla media. La mayor parte de las fracturas son longitudinales, siete en total; hallándose una oblicua.
Alteraciones antrópicas
Sólo se ha constatado una marca de corte (Lám. 10), una muesca con dos puntos de impacto consecutivos sobre un elemento de mesomamífero y 18 restos con alteraciones térmicas, entre los que destacan elementos de ciervo, gato montés (Lám. 11), conejo y quelonio (Lám. 12).

Lám. 10. Incisiones sobre costilla de mesomamífero. Fuente: elaboración propia.

Lám. 11. fragmentos de caparazón de quelonio, dos de ellos (a la izquierda y abajo) afectados por la acción del fuego. Fuente: elaboración propia.

Lám. 12. Primera falange de gato montés con signos de quemado. Fuente: elaboración propia.
Alteraciones por otros agentes biológicos
Se han registrados dos huesos con marcas de carnívoro. En concreto, tres surcos sobre metáfisis proximal de un fémur de conejo y una perforación sobre un hueso del carpo de cabra montés (Lám. 13). La actuación por parte de roedores se ha detectado en una costilla de animal de talla media. Las vermiculaciones apenas han incidido sobre el conjunto. Se han encontrado sobre cinco restos.

Lám. 13. Perforación por parte de carnívoro sobre hueso del carpo de cabra montés. Fuente: elaboración propia.
Alteraciones físico-químicas
La mayoría de los elementos muestran un estado medio de conservación. La principal alteración físico-química por la que se ven afectados es el manganeso, en un 58% del total. Se ha encontrado concreción sobre la superficie ósea de un 27% de los elementos, aunque se trata en su mayoría de finas capas. La alteración subaérea la sigue en porcentaje con un 19,63 % lo que habla de diferentes tiempos de enterramiento para algunos huesos. En menor medida se hallan alteraciones por agua en forma de redondeamientos (13%) y pulidos (6%), siendo muy escasos el trampling (5%) y la compactación (4%).
Niveles solutrenses
Patrones de fragmentación y fractura
El material óseo de los niveles solutrenses está muy fragmentado (Fig. 5) con algo más del 70% de los restos por debajo de los tres centímetros de longitud. Los resultados de la medición de la circunferencia de las diáfisis (Fig. 6) muestran que el 88% de los fragmentos diafisarios de macrovertebrados presentan un radio de circunferencia inferior al 25%, conservando el total de ésta tan sólo un 5%. Para los elementos de conejo nos encontramos con un 71% de los fragmentos diafisarios completos y valores relativamente paralelos entre el 50 y menos del 25% de la circunferencia.
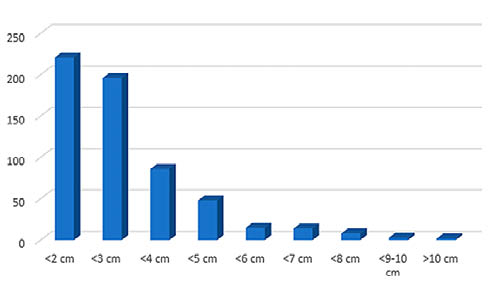
Fig. 5. Patrones de fragmentación en cuanto a longitud de los restos en base al NR para los niveles solutrenses, sin incluir conejo ni erizo.
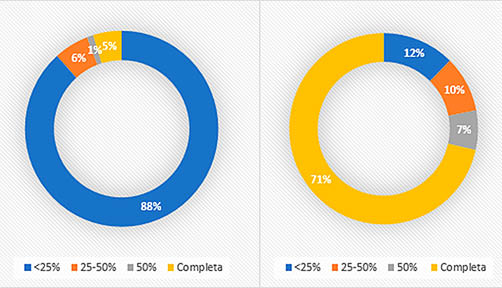
Fig. 6. Grado de circunferencia de los fragmentos diafisarios sin incluir conejo a la derecha y grado de circunferencia de los fragmentos diafisarios del conejo a la izquierda de los niveles solutrenses.
La fracturación de la mayor parte de los huesos largos del conjunto se ha efectuado en fresco (Tab. 10), suponiendo algo más del 60% y sobre planos longitudinales y oblicuos (Tab. 11), entre los cuales los más numerosos son los longitudinales con un 47% del total analizado. Destaca la identificación de numerosos fragmentos óseos fracturados longitudinalmente en fresco para el acceso a la médula, este tipo de actividad se ha visto reflejada sobre todo en las primeras falanges de cabra montés y en los metápodos (Lám. 14).
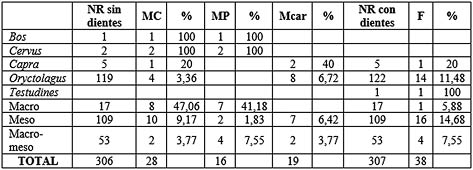
Tab. 10. Tipo de fractura sobre huesos largos en base al NR para los niveles solutrenses. Se han excluido los huesos largos completos (1ª y 2ª falanges, y metápodos de conejo).
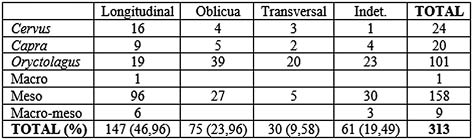
Tab. 11. Morfología de fractura sobre huesos largos en base al NR para los niveles solutrenses.
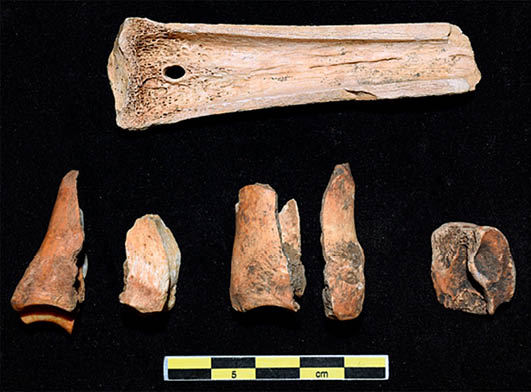
Lám. 14. Muestra de fracturas en fresco longitudinales sobre metacarpo y dos primeras falanges de cabra montés y segunda falange de ciervo para la obtención del tuétano. Fuente: elaboración propia.
Alteraciones antrópicas
La acción humana (Tab. 12) se ve reflejada en los restos óseos analizados de los niveles solutrenses. La frecuencia de marcas de corte no es demasiado alta, pero se encuentran en la mayoría de los taxones. Su presencia en los elementos apendiculares tanto superiores como inferiores plantea un acceso primario a los recursos y un proceso de carnicería en el yacimiento que conlleva un transporte bastante completo de los cuerpos, completado por la presencia de marcas en algunos elementos del esqueleto axial. Se han computado un total de 27 marcas de corte. Las más numerosas corresponden a incisiones practicadas en elementos diafisarios; se constata también la presencia de tajos, aserrados y raspados en menor medida. Estas marcas se han relacionado con las labores de carnicería propias de la acción antrópica, hallándose trazas de desollado, descuartizado, desarticulación (Lám. 15) y descarnación.
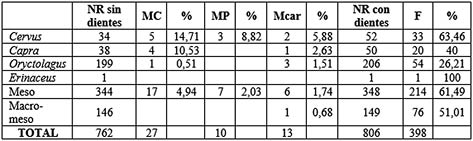
Tab. 12. Resumen de acción antrópica y de carnívoros sobre los taxones documentados en los niveles musterienses. A efectos de porcentajes se han quitado los dientes y clavijas para las marcas de corte, de percusión y de carnívoros. MC: marcas de corte, MP: marcas de percusión, Mcar: marcas de carnívoros, F: fuego.

Lám. 15. Metatarso de cabra montés con incisiones atribuidas a desarticulado. Fuente: elaboración propia.
Las marcas de percusión para los taxones identificados se han encontrado sobre elementos de ciervo, siendo el resto fragmentos óseos de mesomamíferos. Se han hallado estigmas fruto del impacto de percusión, muescas y un cono de percusión.
La evidencia de alteración térmica sobre los restos del conjunto analizado es considerablemente alta, llegando a encontrarse en casi el 50% del total de restos. Se han hallado restos quemados de manera parcial y total (Lám. 16), siendo estos últimos los más numerosos, con tonalidades marrones fundamentalmente.

Lám. 16. Diferentes grados de cremación sobre restos óseos de los niveles del Solutrense evolucionado. Arriba: hemimandíbula izquierda de ciervo. En el medio de izq. a dcha.: diáfisis mesomamífero y dos segundas falanges de ciervo. Debajo de izq. a dcha.: diáfisis de mesomamífero, dos fragmentos de tibia de conejo, fragmento de tejido esponjoso, tres fragmentos de diáfisis de mesomamífero y un fragmento indeterminado de mesomamífero. Fuente: Elaboración propia.
Alteraciones por otros agentes biológicos
Se ha constatado evidencia de 13 marcas de carnívoro (Tab. 12), que implica casi un 2% del total de la muestra, por lo que la incidencia de estos agentes acumuladores es escasa y algunas de sus evidencias se superponen a marcas antrópicas. En su mayoría se trata de impresiones, depresiones, vaciados o surcos (Lám. 17). Son de reducido tamaño por lo que se trataría de pequeños carnívoros que intervendrían de manera secundaria sobre los restos o realizarían aportes de animales como conejos.

Lám. 17. Epífisis proximal de metacarpo de ciervo con signos de vaciado y surcos por parte de carnívoro. Fuente: elaboración propia.
Se ha constatado la presencia de marcas de roedores sobre huesos de conejo y un hueso plano y otro largo de mesomamífero. Las evidencias de vermiculaciones han sido escasas, y las que hay indican un exiguo grado de alteración. Se han contabilizado 97 restos con huellas de este tipo.
Alteraciones físico-químicas
En general, los huesos presentan un relativo buen estado de conservación. Las principales alteraciones de carácter físico-químico que afectan al conjunto están relacionadas con la presencia de agua. La concreción está presente en un 34,85% de la muestra, la mayoría en forma de pequeñas capas. La precipitación de manganeso es bastante notable también, con un peso del 27,46% en el conjunto. El weathering ha sido constatado en un 18,63% del total. Se han comprobado también la existencia de afección por disolución bioquímica en un 14,53%. Además de redondeamientos y pulidos, que corroboran la acción del agua, sumando ambos factores algo más del 10% de la muestra. Parte de los materiales (7,88%) presentan compactación por acción del sedimento y algo más del 5% muestran evidencias de pisoteo o trampling. Sin embargo, se debe tener presente que se trata de sedimentos revueltos por expolio en momentos desconocidos, por lo que su diagénesis se ha podido ver afectada.
DISCUSIÓN Y CONCLUSIONES
En el registro faunístico analizado, nos encontramos con tres conjuntos diferenciados: el de un Musteriense más antiguo que presenta una industria con retoque tipo quina, el del Musteriense con una datación absoluta de 37410±240 BP y con industria lítica de tipo levallois y el del Solutrense evolucionado, con probable interestratificación de un Solutrense superior. Entre ellos parece encontrarse un hiato o ruptura en el que el material arqueológico es prácticamente escaso o nulo, por lo que habrá que esperar a futuras investigaciones para completar esa información.
La interpretación de la secuencia arqueológica de la cueva del Higueral-Guardia es bastante problemática, principalmente por la poca representatividad faunística para cada momento cultural, condicionando que las interpretaciones puedan ser poco definitorias para alguno de esos momentos. Sólo para los niveles solutrenses tenemos algo más de información con el estudio de I. Cáceres (CÁCERES y ANCONETANI 1997; CÁCERES 2003).
La identificación taxonómica ha aportado un patrón que se repite para los tres conjuntos analizados. Ese patrón consiste en un número de especies reducido y la presencia de conejo como la más abundante, seguido por el grupo de los ungulados formado por la cabra montés y el ciervo. En los niveles 8 y 9 musterienses se añade a este grupo el uro, y en los del solutrense el jabalí. Esta parece ser la pauta para la mayoría de los yacimientos con cronologías del Pleistoceno superior del sur peninsular: Higueral de Valleja, (CÁCERES 1997), Gorham’s Cave (RIQUELME et al. 2011), cueva de Nerja (RIQUELME et al. 2006), cueva de los Ojos (TORO y ALMOHALLA 1985), El Pirulejo (RIQUELME 2008), Boquete de Zafarraya (BARROSO et al. 2003) y cueva de Ambrosio (YRAVEDRA 2007).
A esa pauta de conejo y ungulados, se van añadiendo una serie de especies complementarias y representadas en escasa medida. La presencia de restos de carnívoros sólo se ha constatado en el nivel 7 musteriense y en los niveles solutrenses, con el gato montés.
Tras el análisis faunístico se puede establecer que en los tres conjuntos su origen fue fundamentalmente antrópico por todas las evidencias de modificación antrópica (patrones de fractura, marcas de corte, de percusión y alteraciones térmicas) documentadas en el apartado anterior. Esa acción humana se ve plasmada principalmente sobre el grupo de los lagomorfos y los ungulados, uniéndose el consumo de alguna especie de pequeño tamaño como el erizo en los niveles solutrenses o el quelonio en los musterienses. Las aves documentadas se tratan en su mayoría de especies propias de entornos rocosos y cavernosos, probablemente se encuentran allí por muerte natural o aportación de alguna rapaz.
Aunque se ha encontrado actividad por parte de carnívoros, por la escasa presencia de cilindros y la relativa mayor conservación de elementos axiales y apendiculares proximales, más bien estos intervendrían como agentes secundarios y se trataría de pequeños carnívoros que carroñeaban los restos dejados por los humanos, evidenciándose esto en la superposición de las marcas de carnívoros sobre las marcas antrópicas, o aportando pequeñas presas como conejos y otros roedores. Sólo en los niveles 8 y 9 musterienses parece que tuvieron alguna aportación algo mayor, pero no demasiado significativa. A medida que avanza el Paleolítico superior y sobre todo a partir del Solutrense, los yacimientos son ocupados de una manera más continua y sedentaria, lo que explicaría una menor acción de carnívoros sobre los conjuntos óseos. A ello se añade la desaparición en la Península Ibérica de la mayoría de grandes carnívoros (hiena, cuón y pantera) (YRAVEDRA 2001:69).
Sin embargo, es extraño que en los niveles musterienses no se denote momentos de ocupación de carnívoros, como si ocurre en otros yacimientos coetáneos, como la cueva del Boquete de Zafarraya (BARROSO et al. 2003f) o el de la Sima de Constantina (RIQUELME et al. 2017). Aun así, conviene ser cautos en este tipo de interpretaciones, debido al escaso volumen de material musteriense del que disponemos hasta el momento.
Las rapaces no tuvieron una influencia notable, puesto que apenas se han encontrado marcas de pico y no se han hallado signos egagrópilas. Aunque la presencia de cernícalo común en los niveles solutrenses plantea la posibilidad de que realizaran algún aporte puntual.
Tampoco se han hallado elementos en conexión anatómica ni siquiera en conejos, que indiquen una muerte natural en el lugar. Además, las alteraciones fosildiagenéticas como la abrasión, el pulido o el encharcamiento, no indican que los restos llegaran allí por movimientos del sedimento o modificaciones postdeposicionales.
Las modificaciones más abundantes están relacionadas con la presencia de un curso de agua de escasa fuerza, normal en un ambiente kárstico; así como las concreciones calcáreas y las precipitaciones por óxido de manganeso. El sedimento, salvo por la modificación fruto del expolio producido en el yacimiento, tampoco sufrió sustanciales cambios al no ser numerosas las modificaciones por compactación, trampling o abrasión (redondeamiento y pulido).
Para los patrones de transporte de carcasas y aprovechamiento de recursos, la escasa identificación de huesos de especies de ungulados supone un problema para este tipo de interpretaciones. Aun así, se pueden realizar ciertas inferencias. En los niveles 8 y 9, para los macromamíferos, los huesos largos identificados pertenecen al esqueleto apendicular superior (estilopodios y zeugopodios), lo que indica un transporte diferencial relacionado con las partes que aportan más energía y son más fáciles de transportar. También es lógico por el mayor peso de estas especies, suponiendo un primer procesado en el lugar de captación para su mejor transporte a la cavidad. La presencia de costillas y elementos axiales implica igualmente un transporte de estas porciones al yacimiento para realizar allí las labores de carnicería. Del ciervo y la cabra se encuentran principalmente elementos apendiculares. Sin embargo, atendiendo a los elementos asignados a mesomamíferos indeterminados, este espectro se amplía y se documentan vértebras, costillas y un número mayor de huesos largos, que bien pueden pertenecer a estas especies; lo que supondría un transporte más completo de las carcasas.
El nivel 7 apenas nos permite conocer el acceso antrópico a los animales, puesto que los restos disponibles son menores aún, aunque parecen indicar algo parecido a los niveles 8 y 9.
En los niveles solutrenses se documentan todas las regiones anatómicas del ciervo y la cabra, por lo que el aporte de la carcasa al yacimiento fue completo, lo que nos indica que los ungulados pudieron ser cazados y obtenidos en las proximidades del yacimiento, quizás en un radio de unos 5-8 km como así lo indican otros yacimientos coetáneos y con características similares como el de cueva de Ambrosio (YRAVEDRA 2007). Los escasos restos de jabalí no nos permiten hablar de patrones de transporte y procesado y la presencia de un único resto de talla grande nos impide incidir en el consumo de esta talla en estos momentos. Los restos indeterminados asignados al grupo de mesomamíferos presenta elementos que podrían completar los perfiles del ciervo y la cabra como costillas y vértebras, también presenta elementos de las demás regiones.
Los patrones de aprovechamiento cárnico y procesado de los restos muestran ciertas diferencias entre los niveles solutrenses y los musterienses. Teniendo en cuenta que en los niveles musterienses los restos son menos numerosos, el porcentaje de marcas de corte y de evidencias de fracturación intencional es considerablemente más elevado que para los momentos solutrenses que con más del doble de restos, las evidencias de corte y percusión son iguales en número prácticamente a las del Musteriense. No obstante, en los niveles solutrenses el aprovechamiento medular también fue máximo, al encontrarse fracturas en elementos con escaso aporte energético como las falanges y los metápodos, para el acceso al tuétano.
Por otra parte, en el Solutrense se hace un mayor uso del fuego, reflejado en el elevado porcentaje de restos con alteración térmica; mientras que para los momentos musterienses el porcentaje de restos quemados es muy bajos. Ese elevado porcentaje de alteración térmica en el Solutrense puede tener su origen en una ocupación de los asentamientos más alargada en el tiempo (YRAVEDRA 2001:71).
Tanto en los niveles musterienses como en los solutrenses, a partir de las marcas de corte, se ha constatado una completa labor de carnicería y procesado sobre los restos, detectándose para el Musteriense evidencias de desollado, desarticulado, descarnado, evisceración y limpieza del periostio. Mientras que para el Solutrense se han constatado procesos relacionados con el desollado, descuartizado, desarticulado y descarnado.
La gran mayoría de restos de conejo indican que fueron consumidos por el hombre, ya que las marcas de carnicería y afección del fuego son evidentes en los tres conjuntos. Aunque no se descarta que parte de sus restos constituyeron aportación de pequeños carnívoros, por la presencia de marcas de diente y corrosiones en ciertos elementos. Yacimientos como el de Nerja (RIQUELME et al. 2006), Gorham’s Cave (RIQUELME et al 2011), cueva de Ambrosio (YRAVEDRA 2008b) o El Pirulejo (RIQUELME 2008); nos muestran parámetros similares de consumo antrópico para al conejo. Sin embargo, en el caso de Zafarraya (BARROSO et al. 2003c), aunque en los niveles paleolíticos, la presencia de conejo es muy alta, su aportación fue principalmente fruto de alguna especie de rapaz nocturna (probablemente búho real) y algún un mustélido (Barroso et al., 2003c: 239).
No obstante, su pequeño tamaño hace que su aporte cárnico sea escaso, puesto que un solo ciervo o una cabra puede proveer la misma cantidad de carne que un número importante de conejos (YRAVEDRA 2008a: 233), por lo que el hecho de que sea la especie más numerosa no significa que haya una especialización o interés particular sobre la misma. Lo más probable es que supongan un complemento para los momentos de carestía o por su acceso más cercano y fácil sea abundante su caza. Además, conviene no olvidar su presencia relacionada con bioturbaciones, muerte natural u otros agentes biológicos, como los carnívoros.
Respecto a los patrones de muerte y estacionalidad, sólo el material solutrense nos aporta un rango de edades más amplio para los ungulados, teniendo presente que la información es limitada por la fragmentación del material. La selección de la edad en los animales cazados suele estar relacionada con una serie de estrategias cinegéticas condicionadas por el ciclo biológico de las especies. Atendiendo a que es en las estaciones de primavera-verano cuando sería más fácil la captura de las presas por la aparición de los individuos infantiles de los principales ungulados (cabra y ciervo) y la congregación en rebaños de hembras con crías, sería lógico pensar una ocupación temporal durante estos momentos. No obstante, la aparición de individuos juveniles, también de jabalí, ampliaría esa ocupación a periodos otoñales y la presencia de todas las cohortes de edad, siendo más numerosa la adulta y encontrándose individuos seniles, podría significar una ocupación continuada a lo largo del año en el yacimiento, como así ocurre con cueva de Ambrosio (YRAVEDRA 2007 y 2008) y la cueva de Nerja (RIQUELME et al. 2006). Aun así, conviene esperar a obtener más datos para que el NMI se vea aumentado.
Hay que tener en cuenta, en este sentido, que puede ocurrir algo similar al caso de cueva de Ambrosio (YRAVEDRA 2007 y 2008), en el que las cabras, al ser animales con más adaptación a las condiciones climáticas rigurosas y a entornos montañosos, permanecían en los alrededores del yacimiento durante todo el año, suponiendo un recurso continuo disponible. Una hipótesis de trabajo contrastable con futuros estudios que cuenten con más material es que al igual que en cueva de Ambrosio, se produzca una caza diversificada sobre ciervos y cabras en los momentos climáticamente más favorables del año (periodos estivales) y una especialización sobre la cabra en los momentos más crudos (invierno). Aunque no se descartan otras interpretaciones de autores que plantean un comportamiento estacional diferencial en periodos solutrenses (PÉREZ RIPOLL y MARTÍNEZ VALLE 2002).
A partir de las especies animales documentadas se pueden realizar una serie de consideraciones en torno al contexto paleoecológico. Los mamíferos aquí documentados son en su mayoría especies euritermas, es decir, se adaptan bien a diferentes condiciones climáticas. El uro constituye una especie propia de espacios abiertos como praderas y dehesas insertadas en bosques abiertos, con ambientes húmedos y templados. El ciervo necesita un hábitat de bosque denso de especies de árboles caducifolios junto a zonas más abiertas que le proporcionen alimento, está asociado también a climas templados y húmedos, aunque puede aclimatarse a momentos fríos. El gato montés, el conejo y el erizo se suman a este tipo de hábitats de monte mediterráneo con mucha vegetación y matorral (RIQUELME et al. 2011: 171). La cabra montés es una especie rupícola que se mueve por entornos rocosos por encima de los límites del bosque. Mientras que el uro y el ciervo se desplazan buscando condiciones climáticas más benignas, la cabra montés puede permanecer durante buena parte del año, a excepción de los momentos en los que la nieve cubre los pastos de altura (RIQUELME et al. 2011:181).
La paloma y la grajilla son propias también de espacios templados, mediterráneos rocosos y montañosos (BARROSO et al. 2003d:255). El cernícalo tiene capacidad de adaptación a la mayoría zonas climáticas. Tiene preferencia por grandes espacios abiertos con manchas boscosas, tanto en llano como en montaña (BARROSO et al. 2003d:253). La chova piquirroja es una especie cavernícola y xeromontana. La chova piquigualda anida en los espacios rocosos o en las simas, desde 500 a 4000 metros de altitud. Aunque es propia de zonas climáticas mediterráneas, en los momentos fríos del Pleistoceno superior estaría más repartida geográficamente (BARROSO et al. 2003d:258). La golondrina también es una especie de rocas y montañas, extendiéndose por la mayoría de las zonas climáticas. Además, migra en invierno al norte de África, por lo que su presencia aquí indica su aporte en algún momento primaveral o estival (BARROSO et al. 2003d:257).
Los quelonios también pertenecen a climas mediterráneos, aunque relacionadas con una temperatura alta y una insolación estival. Su presencia puede estar relacionada con un periodo estival caluroso (BARROSO et al. 2003b:270). La presencia de rata de agua implica que hubo de haber una fuente de agua constante con abundante vegetación en el área de captación de la cueva (JENNINGS et al. 2009:835).
Se puede considerar que el medio en el que se movían los grupos humanos a lo largo de la secuencia cronológica de la que disponemos, era bastante parecido al actual. Esto es, con unas condiciones de media montaña, con espacios rocosos más altos complementados con un bosque mediterráneo y de matorral, con ciertos claros herbáceos. Ello en un ambiente relativamente templado, lo que concuerda con la datación del nivel 7, que correspondería al estadio isotópico marino OIS3, probablemente a un evento templado del mismo.
Sin duda, uno de los aspectos clave del yacimiento objeto de este trabajo es la presencia de una secuencia que incluye niveles de Paleolítico medio (musterienses) y de Paleolítico superior (solutrenses). A ello es necesario añadir la contribución que supone esta publicación en un panorama científico en el que los estudios zooarqueológicos y tafonómicos en el sur peninsular son más bien escasos. En este sentido, la cueva del Higueral-Guardia presenta un gran potencial para arrojar información sobre las estrategias de subsistencia de los grupos de cazadores-recolectores y la información paleoecológica del Pleistoceno superior, por lo que se espera que los trabajos sobre el yacimiento continúen.
AGRADECIMIENTOS
Resulta imposible no realizar una serie de agradecimientos. A mis directores de Trabajo Fin de Máster: José Antonio Riquelme y Antonio Morgado, por brindarme la oportunidad de hacer un trabajo de estas características y guiarme en su realización. A Javier Baena y al Proyecto Kuretes por la cesión de los materiales y entera disposición. A Alberto Dorado y al Laboratorio de Arqueometría del Departamento de Prehistoria y Arqueología de la Universidad de Granada por el uso de sus instalaciones y equipamiento. Así como al Departamento de Paleontología de la Universidad de Granada por el uso de su colección osteológica. A Antonio Sánchez-Marco por el estudio de la avifauna. Por último, a José Antonio García, Carmen Luzón y Alejandro Beltrán por sus consejos y ayuda.
BIBLIOGRAFÍA
ABRIL LÓPEZ, D. (2012): Contextos arqueológicos de la actividad metalúrgica en el suroeste de la Península Ibérica (III Milenio A.N.E.): la aplicación de análisis zooarqueológicos multivariantes, espaciales y cuantitativos para la explicación de las relaciones sociales, Tesis Doctoral, Universidad de Huelva, Huelva.
AGUAYO DE HOYOS, P.; BARAHONA TEJEDOR, P.; CAPEL MARTÍNEZ, J.; GARRIDO VÍLCHEZ, O. y PADIAL ROBLES, B. (1993): Efectos de la evolución del lapiaz sobre yacimientos situados en calizas: el caso del Cerro de Motillas. Arqueología espacial 16-17, pp. 105-118.
BAENA PREYSLER, J.; MORGADO RODRÍGUEZ, A. y LOZANO RODRÍGUEZ, J.A. (2013): Proyecto Kuretes. Primeras ocupaciones humanas, evolución paleoecológica y climática del Cuaternario de las Béticas occidentales (Serranía de Ronda), CuPAUAM 37-38, pp. 121-138.
BAENA PREYSLER, J.; MORGADO RODRÍGUEZ, A.; LOZANO RODRÍGUEZ, J., TORRES NAVAS, C.; ALCALÁ ORTÍZ, A.; BERMÚDEZ CANO, R.; BERMÚDEZ JIMÉNEZ, F., RUIZ-RUANO COBO, F. (2012): Titanes en el Complejo Motillas. La Secuencia del Pleistoceno Superior de la Cueva del Higueral-Guardia en la Cordillera Bética Occidental. (Proyecto Kuretes), Menga, revista de Prehistoria de Andalucía 3, pp. 106-117.
BARROSO RUIZ, C. (coord.) (2003a): El Pleistoceno superior de la cueva del Boquete de Zafarraya, Junta de Andalucía. Consejería de Cultura, Sevilla.
BARROSO RUIZ, C. y BAILON, S. (2003b): Los anfibios y los reptiles del Pleistoceno superior de la cueva del Boquete de Zafarraya. En El Pleistoceno superior de la cueva del Boquete de Zafarraya (Barroso Ruiz, C., coord.), Junta de Andalucía. Consejería de Cultura, Sevilla, pp. 267-279.
BARROSO RUIZ, C.; DESCLAUX, E.; BAILON, S.; EL GUENNOUNI, K. y ABBASSI, M. (2003c): La fauna de pequeños mamíferos de la cueva del Boquete de Zafarraya. En El Pleistoceno superior de la cueva del Boquete de Zafarraya (Barroso Ruiz, C., coord.), Junta de Andalucía. Consejería de Cultura, Sevilla, pp. 223-250.
BARROSO RUIZ, C.; HERNÁNDEZ CARRASQUILLA, F. y VILETTE, P. (2003d): Los pájaros (aves) del Pleistoceno superior de la cueva del Boquete de Zafarraya (Málaga, España). Estudio comparativo con los yacimientos del Pleistoceno superior antiguo de la cuenca mediterránea. En El Pleistoceno superior de la cueva del Boquete de Zafarraya (Barroso Ruiz, C., coord.), Junta de Andalucía. Consejería de Cultura, Sevilla, pp. 251-266.
BARROSO RUIZ, C.; MARCHI, M. P. de; ABDESSADOK, S.; BAILÓN, S.; DESCLAUX, E.; HERNÁNDEZ CARRAQUILLA, F.; MOIGNE, A. M.; LACOMBAT, F.; LEBRETON, V.; LECERVOISIER, B.; PERRENOUD, C.; RODRÍGUEZ VIDAL, J.; RENAULT-MISKOVSKY, J.; RIQUELME CANTAL, J. A.; VERNET, J. L. y VILETTE, P. (2003e): Contexto paleoecológico, paleoclimático y paleogeográfico de los neandertalenses de la cueva del Boquete de Zafarraya. En El Pleistoceno superior de la cueva del Boquete de Zafarraya (Barroso Ruiz, C., coord.), Junta de Andalucía. Consejería de Cultura, Sevilla, pp. 301-326.
BARROSO RUIZ, C.; RIQUELME CANTAL, J. A.; MOIGNE, A. M. y BANES, L. (2003f): Las faunas de grandes mamíferos del Pleistoceno superior de la cueva del Boquete de Zafarraya. Estudio paleontológico y paleoecológico. En El Pleistoceno superior de la cueva del Boquete de Zafarraya (Barroso Ruiz, C., coord.) Junta de Andalucía. Consejería de Cultura, Sevilla, pp. 169-222.
BEHRENSMEYER, A. K. (1978): Taphonomic and Ecologic Information from Bone Weathering, Paleobiology 4 (2), pp. 150-162.
BINFORD, L. R., (1981): Bones. Ancient Men and Modern Myths, Studies in Archaeology, Academic Press, New York.
BLASCO LÓPEZ, R. (2011): La amplitud de la dieta cárnica en el Pleistoceno medio peninsular: una aproximación a partir de la Cova del Bolomor (Tavernes de la Valldigna, Valencia) y del subnivel TD10-1 de Gran Dolina (Sierra de Atapuerca, Burgos), Tesis doctoral, Universitat Rovira i Virgili, Tarragona.
BLASCO SANCHO, M. F. (1995): Hombres, fieras y presas: estudio arqueozoológico del yacimiento de Paleolítico medio de la Cueva de Gabasa 1 (Huesca), Monografías Arqueológicas, Zaragoza.
BLUMENSCHINE R. J. (1995): Percussion marks, tooth marks and the experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Journal of Human Evolution 29, pp. 21-51.
BLUMENSCHINE, R. J. y SELVAGGIO, M. (1988): Percussion marks on bone surfaces as a new diagnostic of hominid behaviour, Nature 333, pp. 763-765.
BRAIN, C. K. (1969): The contribution of Namib desert Hottentot to understanding of Australopithecus bone accumulations, Scientific Papers in Namibian desert Research Station 32, pp. 1-11.
BUNN, H. T. (1981): Archaeological evidence for meat-eating by Plio-Pleistocene hominids from Koobi Fora and Olduvai Gorge, Nature 291 (5816), pp. 574-577.
BUNN, H. T. (1982): Meat-eating and human evolution: studies on the diet and subsistence patteerns of Plio-Pleistocene hominids in East Africa, University of California, Berkeley.
CÁCERES SÁNCHEZ, I. (1995): Estudio Tafonómico de los Procesos de Formación del Nivel I del Abric Romaní (Capellades, Barcelona): La influencia de la actividad antrópica, Tesis de Licenciatura, Universitat Rovira i Virgili, Tarragona.
CÁCERES SÁNCHEZ, I. (1997): Agentes tafonómicos y económicos de los grupos cazadores-recolectores de la Cueva del Higueral de Sierra Valleja (Cádiz), Revista atlántica-mediterránea de prehistoria y arqueología social 1, pp. 57-76.
CÁCERES SÁNCHEZ, I. y ANCONETANI, P. (1997): Procesos tafonómicos del nivel solutrense de la cueva de Higueral de Motillas (Cádiz), Zephyrus: Revista de prehistoria y arqueología 50, pp. 37-52.
CÁCERES SÁNCHEZ, I. (2003): La Transición de las Sociedades Cazadoras-Recolectoras a Pastoras-Agricultoras en el Mediodía Peninsular a través de los restos óseos. Los modos de vida y de trabajo de las sociedades cazadoras y productoras, BAR Internacional Series 1194, Oxford.
FISHER, J. (1995): Bone surface modifications in zooarchaeology, Journal of Archaeological Method and Theory 2 (1), pp. 7-68.
GARCÍA SOLANO, J.A. (2014): La persistencia de las estrategias de subsistencia de los grupos humanos del Pleistoceno medio, a partir del registro fósil de la cueva del Ángel (Lucena, Córdoba), Tesis doctoral, Universidad de Granada, Granada.
GILES PACHECO, F.; SANTIAGO PÉREZ, A.; GUTIÉRREZ LÓPEZ, J.Mª. y MATA ALMONTE, E. (1997): Las Comunidades del Paleolítico Superior en el extremo sur de Andalucía Occidental. Estado de la cuestión, II Congreso de Arqueología peninsular. Paleolítico y Epipaleolítico. Tomo I, (Balbín Berhmann, R. de y Bueno Ramírez eds.) (Zamora, 1996), pp. 383-404.
JENNINGS, R. P.; GILES PACHECO, F.; BARTON, R. N. E.; COLLCUTT, S. N.; GALE, R.; GLEED-OWEN, C. P.; GUTIÉRREZ LÓPEZ, J. M.; HIGHAM, T. F. G.; PARKER, A.; PRICE, C.; RHODES, E.; SANTIAGO PÉREZ, A.; SCHWENINGER, J. L. y TURNER, E. (2009): New dates and palaeoenvironmental evidence for the Middle to Upper Palaeolithic occupation of Higueral de Valleja Cave, southern Spain, Quaternary Science Reviews 28, pp. 830-839.
MOIGNE, A. M. (1983): Taphonomie des faunes quaternaires de la Caune de l’Arago, Tautavel, Tesis Doctoral, Université Pierre et Marie-Curie Paris-VI, París.
PALES, L. y LAMBERT, Ch. (1971): Atlas Osteologique pour servir à l’identificátion des Mammiféres du Quaternaire, Editions du Centre National de la Recherche Scientifique, París.
PÉREZ RIPOLL, M. y MARTÍNEZ VALLE, R. (2002): La caza, el aprovechamiento de las presas y el comportamiento de las comunidades cazadoras prehistóricas. En De Neandertales a Cromañones. El inicio del poblamiento humano en las tierras valencianas (Villaverde Bonilla, V., ed.), Universidad de Valencia, Valencia, pp. 73-98.
PICKERING, T. R. y EGELAND, C. P. (2006): Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans, Journal of Archaeological Science 33, pp. 459-469.
RIQUELME CANTAL, J. A. (1998): Contribución al estudio arqueofaunístico durante el Neolítico y la Edad del Cobre en las Cordilleras Béticas: el yacimiento arqueológico de Los Castillejos en Las Peñas de los Gitanos, Montefrío (Granada), Tesis Doctoral, Universidad de Granada, Granada.
RIQUELME CANTAL, J. A.; SIMÓN VALLEJO, M. D. y CORTÉS SÁNCHEZ, M. (2006): La fauna de mamíferos del Solutrense en la Cueva de Nerja, Munibe Antropología-Arkeología 57, pp. 255-263.
RIQUELME CANTAL, J. A. (2008): Estudio de los restos óseos de mamíferos de El Pirulejo. Los niveles paleolíticos, Antiquitas 20, pp. 199-212.
RIQUELME CANTAL, J. A.; FINLAYSON, C.; GILES PACHECO, F.; RODRÍGUEZ VIDAL, J.; FINLAYSON, G.; SANTIAGO PÉREZ, A. y Fa, D. (2011): La fauna de mamíferos solutrense de Gorham’s Cave, Gibraltar, Arqueología, Paleontología y Geomorfología del Cuaternario en España: X aniversario del Seminario Francisco Sousa (La Rinconada, Sevilla) (Fernández Caro, J. J. y Baena Escudero, R., coords.), Ayuntamiento de La Rinconada, La Rinconada, pp. 161-178.
RIQUELME CANTAL, J. A.; CARO GÓMEZ, J. A.; ÁLVAREZ GARCÍA, G.; BELTRÁN RUIZ, A.; DELGADO ARCOS, J. A.; SOLER LÓPEZ, A. y CAÑETE GÓMEZ, M. (2017): Presencia de grandes carnívoros cuaternarios en el interior de la cueva de La Sima (Constantina, Sevilla), XI Jornadas de patrimonio histórico y cultural de la provincia de Sevilla. El patrimonio subterráneo (natural o artificial) en la provincia de Sevilla, (Sevilla, 2016), Sevilla, pp. 43-54.
ROSELL ARDÈVOL, J. (2001): Patrons d’aprofitament de les biomasses animals durant el Pleistocè inferior i mig (Sierra de Atapuerca, Burgos) i superior (Abric Romaní, Barcelona), Universitat Rovira i Virgili, Tarragona.
SANTIAGO VÍLCHEZ, J.Mª. (1990): Avance al estudio parietal paleolítico de la Cueva de la Motilla (Cádiz)” Zephyrus: Revista de prehistoria y arqueología 43, pp. 65-76.
SANTIAGO VÍLCHEZ, J.Mª. (2000): “Precisiones en torno al arte paleolítico de las cuevas del cerro de las Motillas. La pintura, Revista de Historia de Jerez 6, pp. 17-36.
SCHMID, E. (1972): Atlas of Animal Bones, Elsevier Publishing Company, Londres
SHIPMAN, P. (1981): Life History of a Fossil. An Introduction to Taphonomy and Paleoecology, Harvard University Press, Cambridge.
SHIPMAN, P. y ROSE, J. (1983): Early hominid hunting, butchering and carcass-processing behaviors: approaches to the fossil record, Journal of Anthropological Archaeology 2, pp. 57-98.
STINER, M. C.; KUHM, S. L.; WEINER, S. y BAR-YOSEF, O. (1995): Differential Burning, Recrystallization, and Fragmentation of Archaeological Bone, Journal of Archaeological Science 22, pp. 223-237.
TORO MOYANO, I. y ALMOHALLA GALLEGO, M. (1985): Un nouveau site du paléolithique supérieur dans le Sud de l’Espagne: le gisement de la Cueva de los Ojos (Cozvíjar, Granada) Note préliminaire, Bulletin de la Société préhistorique française 82 (4), pp.116-119.
TORRES NAVAS, C.; BAENA PREYSLER, J.; MORGADO RODRÍGUEZ, A.; LOZANO RODRÍGUEZ, J.A. y ALCARAZ CASTAÑO, M. (2012): Un enclave solutrense en las cordilleras béticas occidentales: la Cueva del Higueral-Guardia (Cortes de la Frontera, Málaga, España), Espacio, tiempo y forma. Serie I, Prehistoria y arqueología 5, pp. 223-234.
VILLA, P. y MAHIEU, E. (1991): Breaskage patterns of human long bones, Journal of Human Evolution 20, pp. 1-22.
YRAVEDRA SAINZ DE LOS TERREROS, J. (2001): La subsistencia en el Pleistoceno superior en la Península Ibérica: problemas tafonómicos y zooarqueológicos, Espacio, tiempo y forma. Serie I, Prehistoria y arqueología 14, pp. 47-78.
YRAVEDRA SAINZ DE LOS TERREROS, J. (2006): Tafonomía aplicada a la zooarqueología, Universidad Nacional de Educación a Distancia (UNED), Madrid.
YRAVEDRA SAINZ DE LOS TERREROS, J. (2007): Zooarqueología y tafonomía en un yacimiento solutrense del suroeste de la Península Ibérica: La Cueva Ambrosio, Saguntum: Papeles del Laboratorio de Arqueología de Valencia 39, pp. 65-83.
YRAVEDRA SAINZ DE LOS TERREROS, J. (2008a): Especialización y estacionalidad en el Paleolítico superior de La Cueva de Ambrosio, Espacio, tiempo y forma. Serie I, Prehistoria y arqueología 1, pp. 227-238.
YRAVEDRA SAINZ DE LOS TERREROS, J. (2008b): Los lagomorfos como recursos alimenticios en Cueva Ambrosio (Almería, España), Zephyrus: Revista de prehistoria y arqueología 62, pp. 81-99.
