
APROXIMACIÓN AL POBLAMIENTO DEL GÉNERO HOMO EN ANDALUCÍA Y GIBRALTAR: LOS RESTOS ÓSEOS HUMANOS DESDE HOMO SP. HASTA HOMO NEANDERTHALENSIS
AN APPROACH TO THE POPULATION OF THE GENUS HOMO IN ANDALUSIA AND GIBRALTAR: THE HUMAN BONE REMAINS FROM HOMO SP. TO HOMO NEANDERTHALENSIS
Alejandro LEÓN CRISTÓBAL*
Resumen
En este trabajo se muestran los yacimientos del sur peninsular en los que se han hallado restos óseos humanos. El límite cronológico establecido discurre desde los primeros homininos que poblaron el territorio, hasta la llegada de los humanos anatómicamente modernos. El límite geográfico, se acota con los actuales territorios de Andalucía y Gibraltar. Los restos hallados son la evidencia primera de que ambas regiones fueron intensamente pobladas durante la prehistoria antigua.
Palabras clave
Homininos, Sur peninsular, Restos óseos, Poblamiento, Prehistoria antigua.
Abstract
This paper shows the sites in the south of the Iberian Peninsula where human bone remains have been found. The established chronological limit runs from the first hominins population of that the territory to the arrival of the anatomically modern humans. The geographic area includes the current territories of Andalusia and Gibraltar. The remains found are the first evidence that both regions were intensely populated during ancient prehistory.
Keywords
Hominins, Southern Peninsula, Bone remains, Population, Ancient prehistory.
INTRODUCCIÓN. TAXONES HUMANOS ANTIGUOS EN ANDALUCÍA Y GIBRALTAR
Por muchas razones, algunas de índole científica y otras de carácter identitario, el poblamiento del continente europeo emerge como uno de los principales objetivos a la hora de esclarecer el proceso de expansión que llevaron a cabo los grupos humanos desde su primera salida del continente africano (SALA-RAMOS et al. 2011). Esta cuestión todavía no resuelta, tiene una gran influencia para el conocimiento que se posee sobre el continente euroasiático, lugar con nichos ecológicos distintos y diferenciados (DENNELL y ROEBROEKS 2005; BELMAKER 2010) que pese a ello fueron ocupados por nuestros antepasados.
HOMO SP.
Orce
La zona arqueológica Cuenca de Orce posee unas características excepcionales que lo encumbran como una de las más importantes áreas o regiones de la península ibérica. Los yacimientos que conforman la denominada “Zona Arqueológica Cuenca de Orce” poseen una gran importancia a nivel arqueológico y paleontológico, situándose en la llamada cuenca de Guadix-Baza, la cual comenzó a ser investigada desde los años 60, y en mayor medida, en los años 70. Guadix-Baza se encontraría caracterizada durante el Pleistoceno Inferior por un entorno fluvio-lacustre, de gran biodiversidad y clima favorable, que habría permitido el relleno de esta por los sedimentos procedentes del entorno (VILLALOBOS-MEGÍA y BRAGA-ALARCÓN 2003). Estas características ecológicas habrían sido fundamentales para la presencia de diferentes especies biológicas, entre las cuales sin duda habría estado presente el género Homo.
Parece coherente introducir en el presente apartado sobre restos humanos de Orce, la presencia de un resto, inicialmente atribuido a Homo, y que fue hallado en el yacimiento de Venta Micena. Este resto óseo, sumamente discutido e icónico por la relevancia que tuvo en su momento, pasó por diversas variaciones en lo que respecta a su denominación. En primer lugar, fue publicado como humano por Josep Gibert, su descubridor, quien insistió en su categorización como resto antropológico. Posteriormente, su discutida “humanidad” fue apoyada por diversos artículos, como el de Gibert y Palmqvist (1995) sobre la complejidad de las suturas craneales que aparentemente presentaba el resto, o el de Borja et al., (1997) sobre la realización de diversos análisis inmunológicos. Sin embargo, esta atribución duró más bien poco, debido a que uno de los autores que había asegurado su “humanidad” en un principio, Palmqvist, se desdijo de los resultados de la publicación anterior (1997). Años más tarde, Martínez-Navarro (2002) publicaría definitivamente que el discutido y emblemático resto se correspondería con fragmentos de los frontales de una hembra de rumiante, una conclusión que invalidó su correspondencia con el género Homo.
Entre todos los restos hallados presumiblemente humanos, destaca la evidencia, quizás más clara (MONTES-GUTIÉRREZ 2013), de un fragmento de molar inferior deciduo (Fig. 1), atribuido a H. sp. y que apareció en Barranco León 5 con una cronología aproximada de 1.4 millones de años. Además de este fragmento, se hallaría posteriormente un molar completo a escasos metros del anterior y con una cierta semejanza, por lo que serían dos los molares encontrados en el yacimiento de Barranco León (Orce), como se discutirá a continuación.
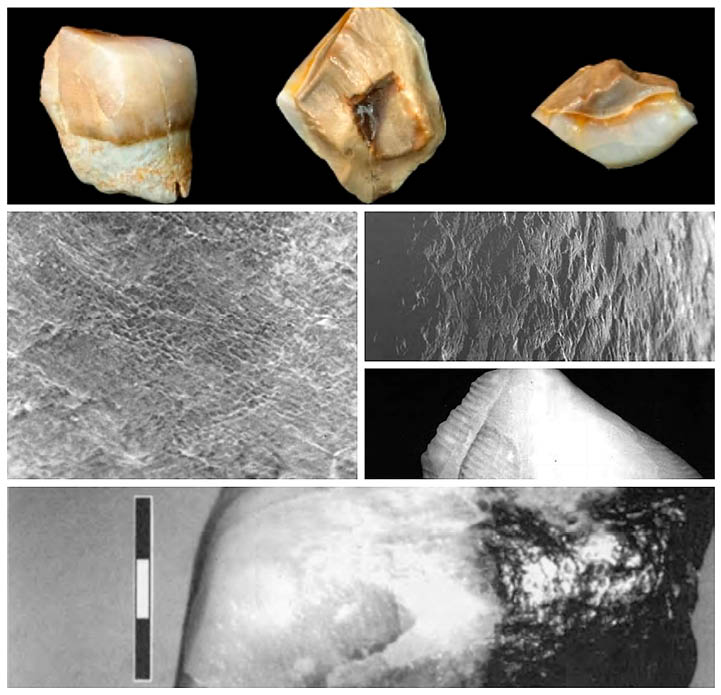
Fig. 1. BL5-0. Molar hallado en Barranco León (Orce). Elaboración propia a partir de Gibert et al., 1999.
Respecto al primero de ellos, Josep Gibert publicaría un artículo (GIBERT et al. 1999) en el que se realizaban varios análisis dentales con el objetivo de demostrar que los fragmentos pertenecían efectivamente a un diente humano. Entre estos análisis se realizaron algunos del esmalte, algunos relacionados con las estrías de Retzius, para comprobar el crecimiento y edad del posible individuo, la presencia de periquimatas, el grosor del esmalte en comparación con otros mamíferos, etc… Gibert y su equipo establecieron que se trataba de un molar inferior deciduo, de un individuo joven y que nada tendría que ver con otros mamíferos con los que estableció los análisis comparativos, como hienas, cercopitécidos, felinos, cánidos, úrsidos, suidos o artiodáctilos. Sin embargo, este hallazgo fue catalogado como paleontológico por Paul Palmqvist, quien lo consideró como perteneciente a la especie Hippopotamus antiquus, un mamífero cuyos restos se encontraban presentes en Venta Micena y Barranco León, rechazando por tanto toda posibilidad de que perteneciera al género Homo. Este fragmento sería revisado además en un artículo científico, más tarde, por Ribot et al., (2015) como se indicará posteriormente.
El segundo de los dientes hallados en Barranco León, fue un primer molar inferior izq. (M1), esta vez mucho más completo que el anterior y publicado en el año 2013 por Toro-Moyano et al. Los análisis realizados mostraron algunas características sumamente relevantes para su estudio, entre ellas el nivel de conservación de las raíces, las cuales se encontraban prácticamente ausentes debido a una más que probable reabsorción, un hecho que ha sugerido que la pérdida del diente fue ante-mórtem (TORO-MOYANO et al. 2013). Los análisis dentales revelaron asimismo la presencia escalonada en tamaño de las cúspides dentarias, siendo la más grande el protocónido, seguido del metacónido, el hipocónido y el entocónido. Este tipo de características dentales serían comparadas con otros datos establecidos en fósiles diferentes, mostrándose que el diente compartía ciertas características generales halladas en el género Homo, pero también algunas muy prometedoras con parántropos y australopitecos. Las conclusiones determinarían que el molar inferior izquierdo, junto con las evidencias de útiles líticos hallados en Barranco León (BARSKY et al. 2014), confirmarían la presencia humana en el oeste europeo por parte de Homo más allá del medio millón de años que se establecía en décadas anteriores y después de la primera salida de África (SALA-RAMOS et al. 2014, 2016), un hecho que se constataría con las evidencias halladas en el yacimiento de Dmanisi (República de Georgia). Más tarde, se concretarían diversos detalles acerca del molar, como por ejemplo que pertenecería a una niña o niño de unos diez años aproximadamente (TORO-MOYANO et al. 2013:7).
Como ya se ha indicado anteriormente, ambos restos aparecieron muy próximos entre sí. Esta relación espacial planteó ciertos problemas y dudas a la hora de su referenciación estratigráfica para algunos autores, entre ellos Arribas y Palmqvist (2002). Estas dudas incidieron en la relación de los restos con la industria musteriense asociada. Sin embargo, diversos artículos posteriores invalidaron esta relación y demostraron que la tecnología presente en el estrato donde fueron hallados era totalmente olduvayense (BARSKY et al. 2010; TORO-MOYANO et al. 2011; TITTON et al. 2020).
Si se compara este diente con BL5-0 se pueden observar algunas características comunes que parecen evidenciar una relación dental incluso espacial, pudiéndose tratar uno como consiguiente del otro.
Según Ribot et al., (2015), ambos dientes tienen el mismo grado de desgaste en el esmalte, así como una morfología en tamaño y dimensiones prácticamente semejante. Además, ambos dientes, observándose de manera conjunta, parecen pertenecer al mismo individuo y dada la distancia espacial entre uno y otro (no más de nueve metros), todo parece indicar que así fue. Si se comprobara que ambos dientes ocupaban un espacio contiguo en la mandíbula, sería bastante probable que pertenecieran a la desarticulación de un individuo fallecido y no a un evento ante-mortem, como así sugería Toro-Moyano et al., (RIBOT et al. 2015). De comprobarse esta cuestión, serían dos los dientes hallados en Barranco León y adscritos a H. sp. lo cual indicaría que más restos óseos humanos podrían hallarse en este yacimiento (SALA-RAMOS et al. 2014).
HOMO HEIDELBERGENSIS
Antigua Cantera de Ágata (Monda)
En el yacimiento de Antigua Cantera de Ágata, localizado en el municipio andaluz de Monda (Málaga), se halló uno de los restos óseos humanos más antiguos de Andalucía. En una pequeña cueva cercana a diversas canteras de áridos, se halló un resto óseo de reducido tamaño cuya adscripción al género Homo no parecía plantear dudas.
El hueso, un temporal izquierdo, presenta una estructura compleja, irregular y que comprende tres funciones de gran importancia: forma parte de la pared lateral y de la base del cráneo; contiene el órgano de la audición; y contiene el órgano del equilibrio, características que lo convierten en un resto de gran valor e interés para el conocimiento de la especie a la que perteneció (Fig. 2).

Fig. 2. Hueso temporal izquierdo de Monda (Málaga). Tomado de: Urquiza et al., 2005.
La datación aproximada se estimó en el Pleistoceno Medio, en torno a los 300.000-500.000 años de antigüedad. Este contexto paleogeográfico sirvió para adscribir la condición de H. heidelbergensis al hueso temporal izquierdo hallado, una especie que habría surgido hace más de 600.000 años y cuyo holotipo, conocido como la mandíbula de Mauer, se encontró en Heidelberg (Alemania).
El estudio del hueso temporal izquierdo se llevó a cabo en la facultad de Medicina de la Universidad de Granada, lugar donde se analizaron las posibles patologías que pudiera poseer el individuo en el momento de su muerte. En primer lugar, se llevó a cabo un escaneado 3D de la superficie del hueso con el objetivo de obtener imágenes de alta resolución para posibles futuros estudios. La presencia de la cavidad auditiva en el hueso temporal permitió observar los conductos auditivos para plantear que efectivamente se trataba de un hominino bípedo, como era H. heidelbergensis (URQUIZA et al. 2005). Tras el escaneado, se compararon los resultados con otros especímenes y se realizaron análisis más profundos del oído medio y de las partes más internas del aparato auditivo. Además, se realizó una reconstrucción en 3D de estas precisas partes, con el objetivo de averiguar el volumen total de las cavidades interiores. Por último, los resultados arrojaron datos prometedores para el conocimiento de esta especie, puesto que mostraron que, en líneas generales, las dimensiones del aparato auditivo y de las cavidades sonoras eran superiores al resto de especímenes consultados, si bien, aunque estas características resultaron diferentes en comparación al aspecto exterior, el contenido del aparato sonoro era prácticamente similar, a excepción de las cavidades timpánicas (URQUIZA et al. 2005:461). Este último hecho parece estar relacionado con algunas otras inferencias presentes en esta especie, como una mandíbula voluminosa y fuerte, con una poderosa musculatura (TORO-IBACACHE et al. 2015; GODINHO et al. 2018) y que podría mostrar las características necesarias para un consumo de carne habitual e importante en la dieta de estos homininos.
HOMO NEANDERTHALENSIS-NEANDERTALES
Cueva Belda (Málaga)
Cueva Belda fue excavada por primera vez en el año 1979. La cueva había sufrido de notables expolios y remociones de tierra, con lo que resultó ciertamente complicada la metodología a emplear y el espacio a excavar. Una vez que metodológicamente se instalaron las pautas a seguir, los resultados fueron rápidos y ciertamente precisos. Tras una ligera capa arcillosa que resultó ser el suelo de habitación, comenzaron a salir materiales asociados a una industria de tipo musteriense, junto con los restos de lo que parecía un pequeño hogar. Una treintena de piezas resultaron de la excavación de este nivel estratigráfico, si bien casi una docena fueron desechadas por no contener ningún tipo de interés arqueológico o por ser únicamente virutas o microlascas (RAMOS-FERNÁNDEZ 1982:17). Este nivel musteriense, muy rico en datos arqueológicos se encontraba prácticamente sumido en un reducido espacio que seguramente sirvió de parapeto para sus moradores. La mayoría del utillaje lítico hallado, se encontró muy patinado, fundamentalmente las piezas realizadas en sílex, y realizado en más de la mitad del porcentaje hallado en caliza dura.
En lo que respecta a los restos óseos humanos hallados en esta localización, en Cueva Belda se limitan única y exclusivamente a un occipital casi completo de lo que parece un individuo neandertal que se adscribiría a la industria musteriense localizada en el interior de la propia cavidad (RAMOS-FERNÁNDEZ 1982:23). Si bien el estado de conservación del resto óseo es bueno, el hueso humano presenta un aspecto más bien arcaico y las múltiples esquirlas de hueso halladas junto al occipital no son suficiente para una valoración antropológica completa.
Además, a día de hoy no existe un estudio específico de este resto óseo puesto que se encuentra en paradero desconocido, por lo que hasta que no proceda, lo único que queda es asignarlo temporalmente a un individuo neandertal o a un humano anatómicamente moderno, pero en este último caso, esto desembocaría en la necesidad de una convivencia temporal entre neandertales y humanos anatómicamente modernos o de un reemplazamiento de estos últimos sobre las poblaciones anteriores, un hecho que únicamente a partir de un resto óseo no analizado resulta cuando menos en una afirmación ambiciosa.
La Cueva de la Carigüela (Granada)
La cueva de la Carigüela se encuentra próxima a la localidad de Píñar, aproximadamente a unos 45 km al noreste de Granada. En el entorno de la zona, se han encontrado también algunas cavidades que rodean el paraje y que al igual que Carigüela permiten controlar desde la seguridad de la altitud todo el territorio. La ubicación de Carigüela, al norte de Sierra Nevada y próxima al curso del río Píñar, resulta idónea desde un punto de vista geográfico para el acceso al valle que se abre hasta el monte La Sierra, a unos 12 km al sur de Jaén.
Al comienzo de la campaña de excavación liderada por Obermaier y Spahni, se descubrió una gran continuidad estratigráfica (Fig. 3) que abarcaba desde finales del Pleistoceno Medio hasta prácticamente la Edad del Bronce (VILLAR-CALVO 1998). Al poco tiempo se hallaron restos óseos humanos, restos que sin lugar a dudas se correspondían con H. neanderthalensis y humanos anatómicamente modernos.
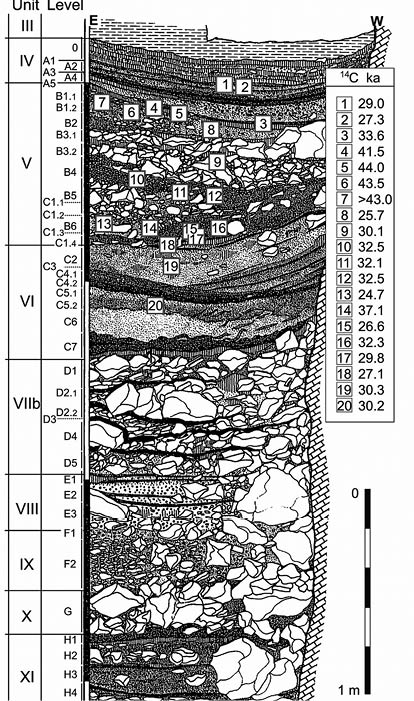
Fig. 3. Corte estratigráfico y ubicación de las unidades VIII, VI, V y IV de la Cueva de la Carigüela. Tomado de: Carrión et al., 2019.
En lo que respecta a los huesos humanos atribuidos a neandertales, estos se correspondían con: dos fragmentos de parietales de pequeño tamaño y que posiblemente pertenecieron a individuos adultos; un frontal infantil; seis dientes de un posible niño neandertal; y tres molares de un más que probable individuo adulto. Todos estos restos esqueléticos y dentales procedían de las unidades VIII, VI, V y IV en su parte más inferior (CARRIÓN et al. 1998:319; CARRIÓN et al. 2019).
En lo que respecta a los dos fragmentos de parietales, tras diversos análisis, pudo observarse que ambos pertenecerían a individuos masculinos y adultos. El primero de ellos fue hallado por Spahni en abril de 1955 en el nivel VI, a unos 1.65 m de profundidad, según indica García-Sánchez (1960).
Este resto óseo se trataba de un parietal izquierdo, de pequeñas dimensiones y con forma ovoidea apuntada. Según García-Sánchez, el espesor del hueso era de considerables dimensiones, muy semejante a los hallados en Cova Negra, pero superado por otros de carácter europeo como los de Spy II, La Chapelle-aux-saints, Krapina o Spy I (DE-LUMLEY 1973).
Una de las características mejor definidas para su adscripción a la especie neandertal era la presencia de una sutura lambdoidea poco compleja (GARCÍA-SÁNCHEZ 1960:24), pero también se observaron otras relevantes cuestiones como una línea temporal desarrollada, o una sinóstosis (fusión de los huesos conforme avanza la edad) avanzada en su parte interna, un hecho que ha llevado a plantear una edad aproximada de entre 35-40 años para este individuo. Hay que destacar también la presencia del llamado hueso wormiano (DE-LUMLEY 1973:304), sobre el cual existe otro de dimensiones reducidas que induce a pensar que se trata de una característica ósea común en H. neanderthalensis, como así señala García-Sánchez (1960).
Sin embargo, estudios recientes han evidenciado algunas patologías congénitas que poseían los neandertales y que pueden estar relacionadas con la endogamia y su desaparición mayoritaria (VAESEN et al. 2019), al igual que la variabilidad presente en el hueso wormiano, un hecho que también lleva a plantear esta posibilidad. La cronología estimada para este fragmento óseo se encuentra en torno a los 21.500 años, momento que se encuentra bien definido paleoambientalmente por las series de polen obtenidas del yacimiento. Este individuo se desplazaría por un paisaje de bosques abiertos de pinos, con presencia de especies mediterráneas que se expandirían exponencialmente por el territorio, con una fauna cada vez más mediterránea y abundante.
El segundo de los fragmentos de parietal, pertenece también a un individuo masculino y es de orientación izquierdo. Su espesura es superior al parietal I y a todos los ejemplos expuestos anteriormente. Su conservación es bastante buena y de ella se ha podido observar una sutura simple como el anterior resto óseo, hecho que confirma de nuevo su adscripción a H. neanderthalensis y una sinóstosis apenas iniciada, característica que ha llevado a plantear una edad aproximada de entre 20 y 25 años (GARCÍA-SÁNCHEZ 1960). Debido a la reducida talla del parietal resulta complicado establecer su posición craneana con exactitud (DE-LUMLEY 1973:306). La cronología que se mantiene para este resto óseo es superior al parietal I, rondando aproximadamente los 30.000 años. Este momento cronológico también está bien definido palinológicamente en las series de Carigüela por un paisaje abierto, con Pinus dispersos y con especies arbustivas que se extienden rápidamente en los momentos más secos y templados.
En lo que respecta al frontal infantil, en 1955, mientras Spahni llevaba a cabo una campaña de excavaciones en la cueva de la Carigüela, encontró una serie de restos óseos humanos que poco tiempo después se asignarían a H. neanderthalensis, uno de los cuales correspondería precisamente a este frontal neandertal (CARRIÓN et al. 1998, 2019; GARRALDA 2005) (Fig. 4).

Fig. 4. Frontal infantil neandertal de Carigüela. Tomado de: Elaboración propia.
Según García-Sánchez (1960), el primero en estudiar este resto óseo, la pieza se encontraba dividida en dos pequeños fragmentos, faltando principalmente la parte superior de estos. Además, la sutura coronal solo se encontraría conservada en la parte más inferior del frontal, una cuestión que provocaría un pequeño hiato entre ambas partes óseas (JIMÉNEZ-ARENAS et al. 2019). Tras el estudio de García-Sánchez, la pieza fue reconstruida físicamente y en la actualidad se encuentran ambas partes unidas.
Se puede decir que el estado de conservación del frontal es bueno, teniendo en cuenta que se han conservado incluso algunos restos de las cavidades nasales en conexión anatómica. Sin embargo, se han perdido parte de la cara y de la nariz, que originalmente se encontraban presentes. Los últimos estudios realizados sobre el frontal muestran que únicamente se conserva el 50% de la superficie endocraneal total de lo que debería ser un frontal infantil (JIMÉNEZ-ARENAS 2019:91). Mediante una serie de reconstrucciones virtuales, se ha podido corroborar lo que algunos autores sospechaban y es que el estado de conservación del hueso y especialmente el de la parte izquierda, se encuentra mucho más afectado debido a deformaciones postmortem que habrían incidido profusamente en la pieza.
En lo que respecta a la edad, en el estudio que realiza García-Sánchez en 1960, indica que no se aprecia sinóstosis en la tabla interna del frontal, uno de los hechos que llevó a indicar aproximadamente la edad que podría tener el individuo neandertal a la hora de su muerte. Además, existiría una ausencia importante de osificación en la sutura coronal, un hecho que nuevamente remite a hablar de un individuo joven de no demasiada edad. García-Sánchez indica que pese a que la ausencia de más datos como serían dientes u otros elementos con lo que precisar la edad, en comparación con otros frontales infantiles de neandertales, el individuo en cuestión tendría entre aproximadamente 7 y 8 años, y correspondería sexualmente a un varón, debido al desarrollo del torus frontalis (GARCÍA-SÁNCHEZ 1960).
De-Lumley publica su estudio sobre el frontal infantil de Carigüela en 1973. En esta publicación, indica en un primer momento, además de por las razones expuestas anteriormente por García Sánchez, que el individuo, claramente neandertal, no tendría más de 6 años. Sin embargo, va más allá y compara las dimensiones y el grosor del frontal infantil de Píñar, con la estimación de edad que dan otros autores para diferentes frontales infantiles estudiados, como los de Starosèlje, Pech-de-l’Azé, Gibraltar, Skhul o La Quina (DE-LUMLEY 1973:252).
Los recientes estudios publicados, han evidenciado algunas cuestiones de especial relevancia, como por ejemplo que los estudios más antiguos no iban mal orientados con respecto a la edad del individuo neandertal, estimándose esta entre 4 y 6 años. Se ha confirmado además la total adscripción de este individuo con los H. neanderthalensis típicos del Würm, los cuales poseen rasgos morfológicos claros y evidentes que los diferencian de los humanos modernos, no solo por los rasgos físicos sino también por las morfologías cerebrales, claramente diferenciadas incluso al nacer (JIMÉNEZ-ARENAS 2019:95). Los diferentes análisis expuestos, además de las reconstrucciones virtuales realizadas en el frontal infantil de Carigüela han permitido visualizar no solo algunas características morfológicas y dimensiones cerebrales que poseían los neandertales infantiles, sino que además han contribuido de esta manera a entender el crecimiento de las cavidades endocraneales paralelamente al crecimiento del individuo, ciertamente diferenciado del de los humanos modernos (Fig. 5).
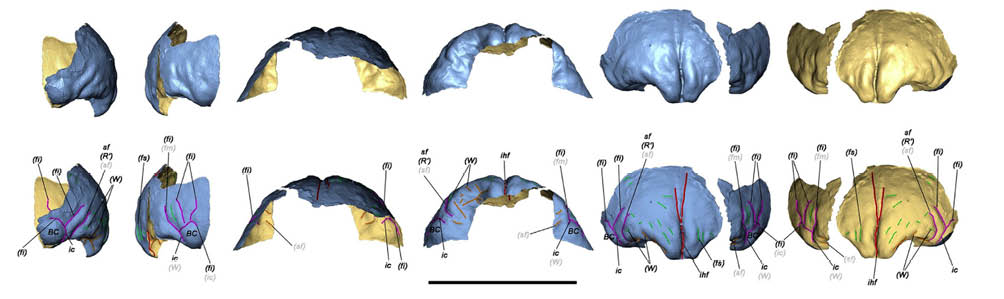
Fig. 5. Análisis y reconstrucciones virtuales realizadas sobre el frontal infantil de Carigüela. Tomado de: Jiménez-Arenas et al., 2019
Además de los dos parietales y del frontal neandertal, se hallaron también seis dientes neandertales, aparentemente de un individuo infantil (CARRIÓN et al. 1998, 2019; GARCÍA-SÁNCHEZ et al. 1994; GARRALDA 2005). En 1969, tras las campañas de excavación de Spanhi, se hallaron en la sala 1 de la cueva de la Carigüela seis dientes que por su tamaño y por el conjunto de restos musterienses con que aparecieron, fueron asignados como neandertales. En este mismo nivel, cerca de las industrias musterienses y de los dientes, se encontraron abundantes restos de fauna típicamente cazada por H. neanderthalensis, como caballos, ciervos, tortugas, bóvidos (STRINGER et al. 2008; NABAIS y ZILHÃO 2019; FIORENZA et al. 2020; ZILHÃO et al. 2020), etc… En 1994 se llevaría a cabo el estudio de estos seis dientes de neandertales (GARCÍA-SÁNCHEZ et al. 1994) hallados en la cueva de la Carigüela, pero no sin ciertos problemas cronoestratigráficos, debido a que la posición de los dientes en relación a la secuencia del relleno de la gruta no está demasiado clara (GARRALDA 2005).
Este estudio de 1994, permitiría la identificación de los diferentes dientes hallados en la cueva, mostrando la presencia de cuatro dientes permanentes, de los cuales dos serían incisivos (I), y los otros dos un canino (C sup.) y un primer premolar (Pr1) superior, todos ellos izquierdos. Los dos dientes restantes, ambos también izquierdos, serían deciduos, es decir, dientes de leche, y se corresponderían con un canino (C sup.) y un segundo molar superior (M2). La identificación morfológica realizada sobre los dientes mostró que todos ellos pertenecerían al mismo individuo y que este tendría en torno a unos 7 años de edad. Uno de los criterios que se utilizó para evidenciar las anteriores afirmaciones, sería que las líneas de hipoplasia localizadas en el esmalte de los dientes se corresponderían con los adyacentes (GARCÍA-SÁNCHEZ et al. 1994).
Estas líneas de hipoplasia se producen por una enfermedad de los dientes derivada de una infección, de una malnutrición o de una afección de carácter medio-grave, generalmente en las etapas de niñez y juventud, momentos en los que se están desarrollando los dientes. Es por ello frecuente que en periodos en los que el niño no ha comido adecuadamente o ha habido una carencia de leche materna por parte de la madre, se hayan originado estas líneas en el esmalte que implican un desarrollo inadecuado de este y de su dentadura. Este tipo de afección se refleja realmente bien en los dientes y permite obtener un alto grado de información acerca de las patologías que presentaba este individuo, así como estimar su edad (RODRÍGUEZ-CUENCA 2003).
Además de estas hipoplasias, también se localizaron algunos defectos de amelogénesis que habrían afectado de manera importante a los dientes permanentes y fundamentalmente al esmalte de estos. Gracias a estos defectos genéticos se ha podido averiguar que en torno a los 2-4 años de edad, se produjeron diversas situaciones de estrés, posiblemente alimenticio y relacionado con el destete, que desembocaron en el surgimiento de este tipo de enfermedades dentales (GARRALDA 2005:303).
Al estimarse que los seis dientes pertenecen a un mismo individuo, se agruparon todos ellos bajo la denominación de Píñar 7 para facilitar su estudio. El primero de ellos (Fig. 6), un C izq. sup. deciduo, posee una raíz robusta y completa que ocupa 2/3 de la altura total del diente. Su predominancia bucal y mesio-lingual y su corona dental desgastada pero caracterizada por una protuberancia lingual sin ningún tipo de bipartición, parecen indicar sin duda alguna que sus atributos se corresponden con neandertales (GARCÍA-SÁNCHEZ et al. 1994).
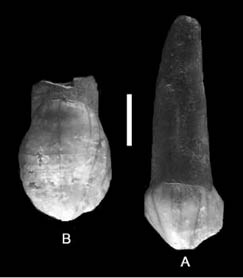
Fig. 6. Canino deciduo y canino definitivo, Carigüela. Tomado de: Garralda, 2005: 304.
El segundo de ellos, otro diente deciduo pero esta vez M2 izq. conserva únicamente la corona. Si bien tiene un alto grado de desgaste en su superficie oclusal, se puede observar la distribución clásica en cuatro cúspides: dos bucales y dos linguales. Según García-Sánchez et al, parece observarse una pequeña cúspide de Carabelli. Esta, no es sino una característica heredable del diente, que genera una cúspide en este, que puede variar en tamaño, y que está hecha sólo de esmalte y dentina, sin ni siquiera presencia de raíz (RODRÍGUEZ-CUENCA 2003). Por último, la corona del diente presenta un cierto estiramiento mesio-bucal, característica que afectaría tanto a neandertales como a los humanos anatómicamente modernos (GARCÍA-SÁNCHEZ et al. 1994:81).
En lo que respecta a los dientes permanentes, cabe comenzar por los dos I izq. (Fig. 7). El primero de ellos, un I izq. lateral, posee una morfología en pala con una ligera desviación en la altura bucal y lingual de la corona. El segundo de los I izq., el central, posee una raíz realmente robusta y una clara presencia de periquimatas a lo largo del esmalte dental (GARRALDA 2005). Las periquimatas son líneas de crecimiento que aparecen en la superficie del esmalte de los dientes y que permiten evaluar cuantitativamente el tiempo de duración y crecimiento de una corona dental.
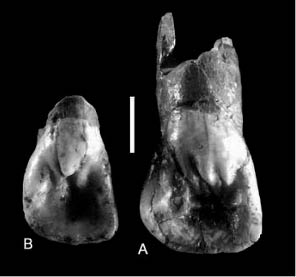
Fig. 7. Incisivo central e incisivo lateral, Carigüela. Tomado de: Garralda, 2005: 304.
El siguiente diente, un C izq. desgastado, y que solo conserva la cara bucal, ha permitido estimar un diámetro mesio-distal de la corona grande, en relación con las dimensiones de los anteriores dientes (GARCÍA-SÁNCHEZ et al. 1994:81).
El último de estos corresponde a un PR1 izq. El estado de conservación de este no es demasiado bueno, faltándole los bordes mesial y distal de la corona, aunque todo parece indicar que se trataba también de un diente de considerables dimensiones.
Por último, hay que tener en cuenta que la publicación de García-Sánchez et al., es el único estudio que hay en referencia a estos dientes presuntamente de neandertales, por lo que no es de extrañar que las conclusiones de su publicación no señalen categóricamente que los dientes pertenecían a H. neanderthalensis y aún menos cuando la cueva de la Carigüela tiene una cronología estratigráfica tan amplia. Si bien la mayoría de las variaciones presentes en el estudio coinciden con estimaciones dadas para los neandertales, hay que tener en cuenta que las dimensiones de los dientes son elevadas en comparación con otros dientes neandertales estudiados, un asunto que contradeciría la hipótesis de que los últimos neandertales mediterráneos sufrieron una reducción en el tamaño de sus dientes (GARCÍA-SÁNCHEZ et al. 1994:82; GARRALDA 1997).
Por último, cabe finalizar el apartado de la Cueva de la Carigüela mencionando el hallazgo de tres dientes de neandertal encontrados en una revisión del material arqueológico obtenido durante las campañas de excavación de los años 1969-1971 y alojados en el museo arqueológico de Granada (JIMÉNEZ-ARENAS et al. 2016). El análisis de estos tres dientes sin publicar ha revelado que se tratarían de tres molares (M1, M2, M3, der.) inferiores derechos (JIMÉNEZ-ARENAS et al. 2016). Estos dientes (Fig. 8) proceden de los niveles XIII a XI, asociados con industria musteriense y con una cronología de más de 46.000 años.
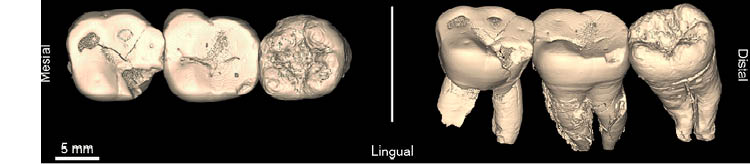
Fig. 8. Tres molares inferiores derechos, Carigüela. Tomado de: Jiménez-Arenas et al., 2016.
Tras los análisis realizados, se confirmó que los tres dientes pertenecerían a un mismo individuo, en este caso un adulto de cierta edad. El primero de los molares (M1 der.) presenta cinco cúspides, al igual que el tercero de ellos (M3 der.), mientras que el segundo molar (M2 der.) únicamente posee cuatro. Sus características dentomorfológicas, junto con rasgos no métricos, se asocian típicamente con la dentición neandertal. Estos estudios añaden más evidencias de la variabilidad presente en los rasgos de los molares entre individuos neandertales.
Cueva del Boquete de Zafarraya (Alcaucín, Málaga)
La Cueva del Boquete de Zafarraya se sitúa próxima al municipio malagueño de Alcaucín. La localización de la cueva permite observar las sierras andaluzas próximas del entorno, hechas de materiales calizos elevados, y que seguramente estarían durante el Paleolítico Medio y Superior cubiertas de nieve.
La situación geográfica y la presencia de numerosos nichos ecológicos son clave para entender la localización de la cueva y la seguridad de este emplazamiento estratégico en su función de asentamiento y hogar de grupos neandertales. Zafarraya se constituye por sus evidencias antropológicas como uno de los grandes yacimientos neandertales del sur peninsular, junto a otros como Carigüela o los yacimientos gibraltareños (RAMOS-MUÑOZ 1998; GARRALDA 2005).
En el año 1979, Cecilio Barroso Ruiz descubrió tras una prospección la cueva del Boquete de Zafarraya, en la cual aparecían en primera instancia numerosos restos óseos de fauna y humanos. Tras lo cual, se llevarían a cabo varias campañas arqueológicas con el objetivo de comprender el tipo de hábitat que habría supuesto la cueva para las poblaciones prehistóricas andaluzas. La obtención de un gran volumen de información acerca de cuestiones faunísticas, paleoclimáticas, botánicas y cronológicas, ha consolidado a Zafarraya como uno de los yacimientos más importantes sobre neandertales de toda Europa.
La cronología aplicada, que se discutirá posteriormente, viene fundamentada principalmente por los análisis realizados sobre muestras de fauna y de carbones presentes en el yacimiento. Los testimonios de hogares basados en cenizas, carbones de madera y huesos quemados reflejarían la presencia humana en la cueva, además de una estrategia de subsistencia ya observada en ocupaciones neandertales.
En lo que respecta a la presencia de restos óseos, cabe indicar en primer lugar que fueron muchos y muy fragmentados los huesos hallados en el interior de la cavidad. Estos aparecieron generalmente junto a otros restos arqueológicos y faunísticos, muchas veces entremezclados y junto a industria musteriense y restos de carbones (GARRALDA 2005; BARROSO-RUIZ et al. 2003). Es por esto que algunos estudios realizados parecen indicar que los restos humanos no provienen de un enterramiento in situ o de sepulturas, como así sucede en otros yacimientos propios del Musteriense, sino que provendrían de niveles de hábitat ricos en desechos culinarios (BARROSO-RUIZ et al. 2003:147). Además, estos niveles son especialmente copiosos en huesos de Capra pirenaica, muy abundante en esta zona de sierra, lo cual parece indicar una clara especialización en la caza de este herbívoro por parte de las sociedades neandertales (BARROSO-RUIZ et al. 1993). La presencia elevada de este animal también tiene un interés añadido, puesto que los homininos de la cueva habrían acumulado intencionadamente restos de estos herbívoros para abastecerse durante el invierno, implicando la práctica de una caza de carácter estacional (MONCLOVA-BOHÓRQUEZ et al. 2012).
Entre las excavaciones de 1981 y 1994, se exhumaron cerca de 55 elementos óseos, correspondientes todos ellos a un número mínimo de nueve individuos y a un máximo de quince. De estos 55 elementos óseos, 25 son unidades anatómicas referidas a 9 dientes aislados, 5 huesos craneales y 12 huesos post-craneales.
En lo que respecta a huesos de neandertal, únicamente son 16 de estas 25 unidades anatómicas los restos óseos que pueden ser asociados con una industria musteriense de forma evidente. Por otro lado, los 9 restantes, parecen asemejarse más con la industria del Paleolítico Superior hallada (BARROSO-RUIZ y DE-LUMLEY 2006).
Los restos únicamente neandertales hallados en Boquete de Zafarraya son los que siguen en la siguiente ilustración (Fig. 9), encontrándose divididos entre los situados en el momento de su excavación en la entrada de la cueva y en el fondo de esta.

Fig. 9. Restos óseos neandertales por zonas de aparición. Tomado de: Barroso-Ruiz et al., 2003: 331.
De entre todos los elementos óseos hallados en la cavidad se han seleccionado para su profundización los siguientes: La mandíbula Zafarraya 2, la mandíbula Zafarraya 4-5-18, los dientes neandertales aislados, la escápula y el húmero Zafarraya 6 y 22 respectivamente, y la tibia Zafarraya 27. Antes de comentar cada uno de ellos, cabe realizar un inciso acerca de la edad de los individuos a la hora de su muerte (BARROSO-RUIZ et al. 2003). A partir de los análisis antropológicos se ha podido constatar la edad aproximada de muerte de unos nueve individuos neandertales, divididos entre niños y adultos. Los dos niños fallecieron en torno a los 14 meses y a los 12 años. Cuatro de los adultos fallecieron entre los 18 y los 20 años, dos entre los 21 y los 30 y del último restante resulta demasiado complicado establecer una edad aproximada. Este análisis de la población neandertal de Zafarraya muestra que la mortalidad más reflejada es la de los adultos-jóvenes, con un 44%, mientras que entre los niños sería del 22% (BARROSO-RUIZ et al. 2003:332).
En lo que respecta a la mandíbula Zafarraya 2 hay que señalar que es el mejor fósil conservado de Zafarraya, de gran tamaño y robustez y muy completa (SÁNCHEZ 1999; GARRALDA 2005; BARROSO-RUIZ y DE-LUMLEY 2006). El estado de conservación es excepcional, puesto que son pocos los ejemplos a nivel europeo que existen de una mandíbula con la práctica totalidad de dientes presentes en los alvéolos, trece de dieciséis, concretamente. Los dientes ausentes, dos I izquierdos y el M3 derecho, se encontrarían presentes en el momento de la muerte del individuo, pero sin embargo se habrían desprendido en un momento post-mortem. Tras la realización de algunos análisis antropológico-forenses, se ha podido constatar que perteneció a una joven neandertal de sexo femenino de entre 25 y 30 años de edad, con una musculatura fuerte y potente, a tenor de la presencia de inserciones musculares marcadas. Pese a que los valores morfométricos de esta mandíbula concuerdan con los establecidos para H. neanderthalensis y no hay lugar a dudas en su adscripción, la ausencia de mentón permite la exclusión de análisis comparativos con el género sapiens (BARROSO-RUIZ et al. 2003:339). Sin embargo, algunos autores sí que han propuesto estas comparativas con otros Homo, como H. erectus, H. sapiens u H. sapiens sapiens (SÁNCHEZ 1999), llegando a la conclusión de que al igual que en la Cueva de la Carigüela, los restos óseos neandertales parecen tener un aspecto mucho más moderno y cercano a los humanos anatómicamente modernos que en otros yacimientos europeos más tardíos. Algunos de estos estudios (RIVERA-ARRIZABALAGA 2008a, 2008b) han señalado que quizás un contacto estrecho con poblaciones sapiens, podría haber desembocado en un proceso de “gracilización” de estas poblaciones neandertales (SÁNCHEZ 1999:135). Sin embargo, aunque esta línea de investigación podría resultar prometedora, sin estudios paleogenómicos no puede ser confirmada.
Los dientes hallados en los alvéolos poseen unas dimensiones grandes, muy similares a las de otros ejemplares neandertales hallados en yacimientos como La Quina 5, Le Moustier o Genay (Francia), aunque se pueden apreciar ciertas diferencias con respecto a los dientes de neandertales mediterráneos, mucho más pequeños estos últimos.
Respecto a la mandíbula Zafarraya 4-5-18, no posee el mismo grado de conservación que la anterior y además se encuentra incompleta (GARRALDA 1997:155). Sin embargo, destaca de ella que se encuentra formada por tres fragmentos óseos mandibulares que se hallaron quemados, perteneciendo a un posible individuo adulto neandertal. Pese a que es menos saliente que la anterior, presenta similitudes muy claras con respecto a Zafarraya 2 (BARROSO-RUIZ et al. 2003:366; GARRALDA 2005:305).
En cuanto a los dientes neandertales aislados, cabe indicar que tanto su morfología como sus dimensiones recuerdan ciertamente a los de H. neanderthalensis, aunque eso sí, a los más robustos (GARRALDA 1997:155). Destaca la presencia de un tubérculo lingual en el incisivo 1 (I1), bien conservado y con morfología en pala. También la presencia de una prominencia basal vestibular es destacable fundamentalmente en los caninos y en los primeros premolares inferiores (PR1, PR2,). Por último, los molares inferiores (M1, M2, M3) parecen tener una corona de cinco cúspides y un taurodontismo poco desarrollado, una característica muy apreciable en las poblaciones neandertales y que consiste en una anomalía que se da en molares y premolares y en la que se produce un aumento de la cámara pulpar muy destacado (RODRÍGUEZ-CUENCA 2003). En términos generales se puede indicar que, junto con las inserciones musculares, estos individuos poseían un aparato masticatorio robusto y fuerte (BARROSO-RUIZ et al. 2003:368).
Continuando con los elementos del miembro superior, cabe destacar la escápula Zafarraya 6 y el húmero Zafarraya 22. El primero de ellos, un pequeño fragmento de escápula hallado en 1990, se encuentra quemado y sería de orientación derecha. Fue hallada junto con otros restos humanos muy fragmentados y quemados, restos faunísticos, cenizas, carbones de madera, etc… La superficie articular no estaba completamente osificada, signo inequívoco de que se trataba de un individuo joven. La estimación aproximada del resto es de unos 18-22 años. En cuanto al húmero Zafarraya 22, hallado en el año 1992, entre niveles musterienses, paleolíticos y neolíticos (GARRALDA 2005), debió de pertenecer a un individuo neandertal robusto, probablemente de sexo femenino. En diversas comparativas realizadas con otros húmeros neandertales, se ha podido comprobar que posee una morfología similar a los hallados en Cova Tossal de la Font (Vilafamés) y Crouzade (Francia) (BARROSO-RUIZ et al. 2003:370). Todo parece indicar que al igual que la escápula Zafarraya 6, perteneció a un individuo joven.
Por último, la llamada tibia Zafarraya 27 se halló en diversos fragmentos revueltos al igual que el húmero anterior entre varios restos óseos quemados. Se conserva únicamente la mitad inferior de la diáfisis y la mitad superior del agujero nutricio, si bien no existe continuidad ósea entre estos dos restos. Los análisis antropológicos han mostrado por su gracilidad que estaría en completa relación con dos fémures, también gráciles, de un individuo neandertal femenino. Como se señala en Barroso-Ruiz et al., (2003: 379), resulta sumamente complicado reconocer rasgos neandertales en restos tan degradados y fragmentados, aunque todo parece indicar que procedían del mismo individuo.
La presencia de estrías de descarnamiento y de posibles fracturas intencionales en algunos huesos hallados como, por ejemplo, las dos mandíbulas o los dos fémures femeninos, además de haberse encontrado indicios de ser exhumados de una pequeña fosa con numerosos restos de carbones de madera, cenizas, restos quemados, etc… parece indicar que fueron sometidos a la acción del fuego (GARRALDA 2005) y según algunos autores a un canibalismo de tipo ritual (BARROSO-RUIZ y DE-LUMLEY 2006; MICHEL et al. 2011:584; COLE 2017).
Cabe finalizar el presente yacimiento indicando un debate relacionado con la cuestión de los últimos neandertales en el sur de la península ibérica. El principal problema de algunos de los yacimientos neandertales del sur peninsular es que casi 2/3 de las dataciones realizadas se utilizaron con técnicas de radiocarbono, una técnica que se ha demostrado como muy sensible a los contaminantes de carbón de temprana edad (WOOD et al. 2013). Esto contaminantes resultan muy complicados de eliminar a la hora de realizar las dataciones usando protocolos de pretratamiento rutinarios. El principal problema es que las nuevas dataciones, empleando métodos de ultrafiltración para purificar el colágeno de los huesos, han mostrado que las cronologías obtenidas por radiocarbono eran al menos 10.000 años más antiguas que estas. Esta polémica comienza a evidenciar que las dataciones radiocarbónicas no son del todo fiables a la hora de establecer cronologías precisas, por lo que hasta que no se empleen otras técnicas que revisen las dataciones del sur peninsular o se apliquen protocolos de pretratamiento rutinario capaces de eliminar la posible contaminación de las muestras, estas han de considerarse como desacertadas y poco precisas (WOOD et al. 2013:2785).
Cueva Horá (Granada)
Cueva Horá, situada próxima al municipio de Darro, en la provincia de Granada, es un buen ejemplo de yacimientos en abrigos rocosos andaluces. Cerca de él se encuentra la sierra de Harana, formada por rocas calizas de origen kárstico. No hay que visualizarlo en este caso como un yacimiento aislado sin ninguna conexión con el resto del territorio, puesto que toda la zona circundante a la sierra de Harana registra una gran ocupación desde el interglacial Riss-Würm y Würm I (HABER-URIARTE 2002; PANIAGUA-PÉREZ 1998), momento en el que las temperaturas son realmente suaves. Posteriormente, según avanza el Würm, el clima se endurece y las poblaciones de homininos se desplazan hacia grietas, abrigos rocosos o cuevas en busca de refugio frente a las condiciones adversas que comienzan a darse en el territorio. Los estudios de la cueva han mostrado un registro sedimentario elevado y muy completo del Pleistoceno (JIMÉNEZ-ARENAS y MARTÍN-LAGOS 2014) y la relativa proximidad con otras zonas de especial relevancia para el poblamiento humano prehistórico como la cueva de la Carigüela, Fuente de la Zarza o Llano de la estación de Huélago, parecen indicar la presencia de poblamiento prehistórico en las inmediaciones de la sierra de Harana (VILLALOBOS-MEGÍA y BRAGA-ALARCÓN 2003).
La cueva, descubierta en el año 1934 por Hugo Obermaier, reveló la presencia de material lítico, todo ello formado por sílex autóctono, probablemente recogido en las inmediaciones de sierra de Harana. Cabe destacar una talla con percutor duro y un retoque con percutor blando (PANIAGUA-PÉREZ 1998).
Tras limpiar la estratigrafía de uno de los niveles más fértiles en industria lítica de este tipo, apareció un resto óseo aparentemente humano, un fragmento de frontal con una conservación muy buena y con una datación de hace aproximadamente 100.000 años (HABER-URIARTE 2002; VILLALOBOS-MEGÍA y BRAGA-ALARCÓN 2003).
Este pequeño fragmento óseo no presenta en principio marcas de cortes ni ningún tipo de alteración, a excepción de una fractura producida por la influencia del terreno sobre el hueso. Este fragmento neurocraneal no permite conocer el sexo del individuo, si bien se ha podido observar a través del estudio antropológico del resto que poseía una pequeña muesca, una cuestión relacionada con el padecimiento de un posible traumatismo craneoencefálico que el individuo sufrió en vida. El estudio del espesor del fragmento craneal ha otorgado ciertas complicaciones a su adscripción, puesto que dentro de la variabilidad característica de las poblaciones neandertales, los rasgos espaciotemporales de la bóveda craneal se asemejan tanto a los humanos anatómicamente modernos como a neandertales, una cuestión que contrastaría con el esquema tradicional de que los neandertales poseían huesos craneales más robustos (HABER-URIARTE 2002:328).
Las comparaciones realizadas con otros frontales infantiles, como el de la cueva de la Carigüela, por ejemplo, han mostrado que el frontal de cueva Horá no se asemeja en dimensiones por lo que no se trataría de un individuo infantil. No obstante, en otras comparaciones realizadas, tampoco se alcanzan los valores específicos para un neandertal adulto en el occidente europeo. Así pues, existen dos posibilidades: En primer lugar, que se trate de un individuo joven, subadulto, masculino o femenino; o, en segundo lugar, que se trate un individuo casi adulto, pero de sexo femenino (HABER-URIARTE 2002).
Gibraltar
Con el gran protagonismo que han tenido los neandertales en los acontecimientos prehistóricos de las últimas décadas, no es de extrañar que uno de los lugares que aportó tan tempranamente un cráneo de neandertal a la comunidad científica sea tan importante para el panorama internacional. Gibraltar, ubicado en el sur de la península ibérica, en la pequeña península de Gibraltar, ha sido protagonista de numerosos restos óseos hallados en algunas de las cavidades que se encuentran próximas al Peñón de Gibraltar, la elevación más alta del territorio.
En el año 1848 se halló en Forbes’ Quarry un cráneo presumiblemente neandertal. La relevancia de este cráneo (Fig. 10) viene marcada por dos curiosidades: en primer lugar, fue hallado antes de que Darwin expusiese su teoría de la evolución; y, en segundo lugar, fue hallado ocho años antes que el cráneo infantil de neandertal hallado en el Valle de Neander (Alemania). Pese a no poseer un buen contexto estratigráfico, cabe señalar que es uno de los cráneos más interesantes hallados en la península. Por desgracia, se encuentra ciertamente incompleto, faltándole casi todo el lado izquierdo y parte de la base craneana (GARRALDA 2005). Su torus supraorbitalis se encuentra bien delimitado y en buenas condiciones, con forma semicircular por encima de las órbitas y con una frente que supera las dimensiones de otros ejemplos de cráneos neandertales. La apariencia del conjunto no hace entrever que se trate de un cráneo de grandes dimensiones, si bien la capacidad craneana se ha estimado en torno a los 1230-1250cc.
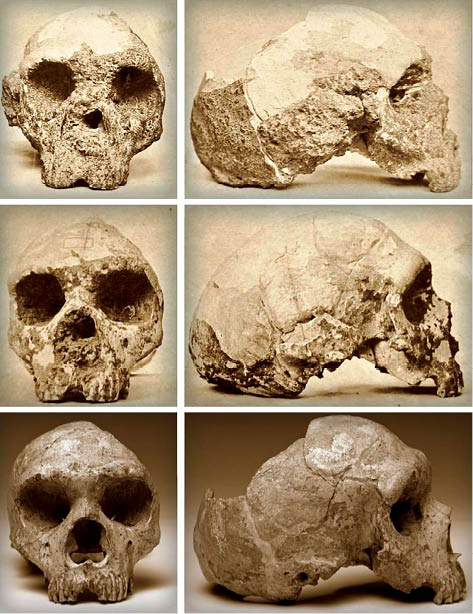
Fig. 10. Cráneo neandertal hallado en Forbes’ Quarry (Gibraltar). Tomado de: Menez, 2018: 105.
Respecto a los dientes que se encontraban en la base alveolar, se puede indicar que están ciertamente abrasionados, conservándose únicamente ocho del total de dientes. Sus coronas apenas son visibles y prácticamente desaparecidas y algunos análisis maxilares han mostrado la presencia de parodontosis generalizada (GARRALDA 2005:306), una fase final de la gingivitis que, si no se trata, culmina con la pérdida inevitable de los dientes afectados, de ahí una posible explicación a la ausencia de varios dientes. Debido a la gracilidad del cráneo, normalmente se ha atribuido a un individuo femenino, si bien algunos autores consideran que podría tratarse de una forma de transición entre los pre-neandertales y los neandertales más antiguos (LACRUZ et al. 2015).
En otra de las cavidades gibraltareñas, Devil’s Tower, aparecieron restos óseos aparentemente humanos de gran valor para la cuestión del poblamiento neandertal. En el nivel atribuido al Musteriense, se encontraron un frontal, un parietal izquierdo, un temporal derecho, un maxilar derecho y una mandíbula, todos ellos conformando conjuntamente un cráneo de lo que parecía un homínido infantil con una datación aproximada de entre 100.000 y 80.000 años (GARROD et al. 1928). Tras los primeros análisis antropológicos, se vio que evidentemente se trataba de un cráneo de un individuo infantil de en torno a unos 4-5 años. La reconstrucción del cráneo permitió observar sus dimensiones y las partes ausentes, fundamentalmente localizadas en la dentición. Resulta interesante la parte del maxilar, puesto que se aprecia la región alveolar muy hinchada debido a que poseía las coronas en formación y los dientes permanentes esperando a emerger. (CARBONELL 1965; GARRALDA 2005). De entre los dientes conservados, destacan dos M izq. deciduos con cuatro cúspides. El M1 izq. presenta además una fosa de Carabelli visible a simple vista. Como se indicaba anteriormente, se aprecian dos dientes permanentes bajo los alvéolos: un molar (M) con cuatro cúspides y un incisivo (I) con una corona de gran tamaño y muy curvada, en forma de “pala” (CARBONELL 1965; GARRALDA 2005). La mandíbula se encuentra incompleta y las inserciones musculares no son demasiado grandes, signo de que pertenecía a un individuo infantil. Cabe indicar por último que se aprecia cierto taurodontismo en los dientes de ambos maxilares, una cuestión que parece repetirse en el caso de otros niños neandertales ya estudiados (BARROSO-RUIZ et al. 2003). De los análisis maxilares se aprecia también que el niño perdió en vida el incisivo inferior derecho, apreciándose las coronas totalmente formadas, pero malposicionadas (GARRALDA 2005), por lo que un posible traumatismo pudo ser el causante de ello.
Un reciente artículo ha permitido verificar que este individuo infantil era un varón, a través del análisis de 36 miligramos de polvo de la parte petrosa del hueso temporal (BOKELMAN et al. 2019). Este mismo trabajo verificó además que el cráneo de adulto hallado en Forbes’ Quarry se trataba de una hembra, como ya algunos autores habían señalado (GARRALDA 2005; LACRUZ et al. 2015), indicando también una cuestión sumamente relevante, y es que Forbes’ Quarry no habría constituido el último refugio de los neandertales antes de su extinción. Esto es así puesto que, tras la recuperación de ADN contaminado por un clima cálido, como era el caso, se ha podido observar que la hembra adulta de neandertal sería genéticamente más parecida a las poblaciones comprendidas entre el 60.000 y el 120.000 BP, que a los últimos neandertales mediterráneos hallados en algunos yacimientos peninsulares (BOKELMAN et al. 2019).
En el año 2017, se halló en otra cavidad cercana, Vanguard Cave, un nuevo resto óseo, concretamente un diente humano. Tras un nuevo proyecto de excavación en el que se buscaba hallar restos óseos de pequeño tamaño de reptiles y roedores, se detectó la presencia de un pequeño canino (C) deciduo (diente de leche), que aparentemente parecía humano. Tras los primeros análisis dentales y antropológicos se ha podido comprobar que el diente correspondería a un individuo infantil que no superaría los cinco años de edad y que vivió hace aproximadamente 50.000 años (GILES-GUZMÁN Y LEE-HOLMES 2018). Las excavaciones continúan avanzando, por lo que se espera que se encuentren más restos de este pequeño niño neandertal que no hayan sido todavía hallados.
La última de las cuevas con presencia humana antigua en el Peñón de Gibraltar es la cueva de Genista, donde en 1865 se halló la presencia de un diente humano y de un cuchillo de sílex, junto al esqueleto de un rinoceronte (GARRALDA 2005). Este diente, fue trasladado al British Museum junto con el resto de colecciones procedentes de Gibraltar. El diente de neandertal permaneció perdido en las bóvedas del museo durante casi 70 años, hasta que el Dr. Hopwood, examinando las colecciones, observó que el diente en cuestión tenía una gran familiaridad con los hallados en Gibraltar. Tras los análisis dentales de la pieza se pudo corroborar su adscripción al género Homo, aunque no se pudo concretar más puesto que poseía unas dimensiones inferiores a las de otros fósiles cercanos, como el M1 de Devil’s Tower. Este diente correspondía a un M1 izq. definitivo que no habría emergido todavía y que por lo tanto tenía un estado de conservación casi perfecto. Los análisis realizados mostraron que pudo posiblemente pertenecer a un niño de unos 5 o 6 años, aunque como ya se ha dicho, resulta dudosa la adscripción neandertal que se le otorgó, debido en gran parte a su descontextualización (GARRALDA 2005:308).
CONCLUSIONES
Como se ha podido observar en el presente artículo, Andalucía posee varios yacimientos con restos óseos humanos que vienen a corroborar la presencia del género Homo en el territorio. Tal y como se ha especificado anteriormente, el presente trabajo limita cronológicamente las especies humanas hasta la desaparición de los últimos neandertales y la llegada a la península ibérica de los llamados humanos anatómicamente modernos. Andalucía y Gibraltar cuentan con registros que van desde la evidencia más factible de la presencia de H. sp. en Orce, como es gracias al hallazgo de, al menos, un fragmento de molar inferior deciduo, hallado en Barranco León; hasta las evidencias más tardías en el tiempo de la presencia de H. neanderthalensis en la pequeña península de Gibraltar.
Estas evidencias del poblamiento de Homo en el sur de la península ibérica han servido para ahondar aún más en las cuestiones del poblamiento y de la evolución humana, generando numerosos estudios y publicaciones de carácter internacional. Gracias a ellas, se ha podido llegar a tener el conocimiento de que el oeste europeo no fue tan tardíamente colonizado por la especie humana como inicialmente se creyó durante décadas. La llegada de Homo a la península ibérica sigue siendo todavía contradictoria en algunos puntos, al igual que el avance de fauna asiática y africana que aparece en el sur del territorio y que parecen establecerse ahí gracias a las óptimas condiciones climáticas que se dan, así como por la mayor abundancia de vida vegetal y de recursos hídricos.
La intensa investigación en Andalucía ha tenido como recompensa la presencia de hallazgos prehistóricos de gran interés para la comunidad científica. El volumen de información extraído de algunos yacimientos, supone un gran paso adelante en el conocimiento de nuestros antepasados que, junto con la colaboración internacional, crece día a día. Yacimientos como Orce o los de Gibraltar han posicionado al sur peninsular como una gran fuente de información acerca de la evolución humana. Andalucía, al igual que Gibraltar, tiene mucho que decir y aportar todavía a la prehistoria europea, y parece evidente que, con la ayuda necesaria, la información que se obtenga en los próximos años sea todavía mayor.
La situación estratégica del sur peninsular entre dos mares y dos continentes ha favorecido la creación de hipótesis de paso a través del Estrecho de Gibraltar, por la que la fauna africana y parte de los homininos que salieron del continente, habrían accedido a Europa a través de Andalucía y Gibraltar. No obstante, hay que tener en cuenta dos hechos para verificar esta hipótesis: que este paso del Estrecho no sería una vía preferente; y que en el yacimiento chino de Shangchen se habría encontrado presencia humana con 2.1 millones de años, lo cual lleva a plantear que la primera dispersión de Homo no se habría realizado a priori por el occidente europeo. Algunos estudios apuntan más hacia una relación estrecha de contactos entre el norte de África y el sur peninsular principalmente durante el Paleolítico Medio.
Por último, cabe señalar que, pese a que únicamente se cuenta en Andalucía y Gibraltar con los presentes restos óseos señalados, resulta factible pensar que sean muchos más los que con el tiempo salgan a la luz. Es vital por ello continuar con la búsqueda de las evidencias prehistóricas que permitan revelar las cuestiones del poblamiento del sur peninsular, y, por ende, de toda la península ibérica.
AGRADECIMIENTOS
Resulta inevitable agradecer en modo alguno el apoyo, los consejos, la dedicación y las buenas palabras de quienes comparten momentos personales con uno mismo en la realización de este tipo de cuestiones académicas. El primer reconocimiento es para mi director de Trabajo de Fin de Máster, Juan Manuel Jiménez Arenas, excelente tanto en las cuestiones personales como en las profesionales y académicas. Su apoyo y su entrega, aun en situaciones de complejidad personal, han sido clave para la realización del trabajo de fin de máster y del presente artículo. A quien debo mi formación prehistórica y mi afición por ella, es a Miguel Ángel Fano Martínez, profesor titular de la Universidad de La Rioja y a quien sin duda aprovecho para agradecer su dedicación y su bondad. Por último, a mis padres, por su apoyo, y por no dejar nunca de creer en mí.
BIBLIOGRAFÍA
ARRIBAS, A., PALMQVIST, P. (2002). The first human dispersal to Europe: remarks on the archaeological and palaeoanthropological record from Orce (Guadix-Baza basin, southeastern Spain). Human Evolution, 1-2, 2002, pp. 55-78.
BARROSO-RUIZ, C., HUBLIN, J.J., MEDINA-LARA, F. (1993). Proyecto: Zafarraya y el reemplazamiento de los neandertales por el hombre moderno – Anatómicamente en Europa Occidental (Objetivos del Proyecto General de Investigación y resumen de los resultados obtenidos en las campañas 1990-1993). Huelva, Junta de Andalucía, Investigaciones arqueológicas en Andalucía (1985-1992), (J.M. Campos Carrasco, F. Nocete Calvo, Eds.), 1993, pp. 229-238.
BARROSO-RUIZ, C., DE LUMLEY, M.A., CAPARRÓS, M., VERDU, L. (2003). Los restos humanos neandertalenses de la Cueva del Boquete de Zafarraya. En: BARROSO-RUIZ, C. (Coord.). El Pleistoceno Superior de la Cueva del Boquete de Zafarraya. Arqueología – Monografías, Junta de Andalucía, Málaga, 2003, pp. 327-385.
BARROSO-RUIZ, C., DE LUMLEY, H. (Eds.). (2006). La grotte du Boquete de Zafarraya. Málaga, Andalousie, Junta de Andalucía. Consej. Cultura, 2006.
BARSKY, D., CELIBERTI, V., CAUCHE, D., GRÉGOIRE, S., LEBÈGUE, F., DE LUMLEY, H., TORO-MOYANO, I. (2010). Raw material discernment and technological aspects of the Barranco León and Fuente Nueva 3 stone assemblages (Orce southern Spain). Quaternary International, 223-224, 2010, pp. 201-219.
BARSKY, D., SALA-RAMOS, D., TORO-MOYANO, I., MENÉNDEZ-GRANDA, L. (2014). La industria lítica en Barranco León y Fuente Nueva 3: Pleistoceno Inferior y Medio de la Cuenca de Guadix-Baza y Orce. Universidad de Burgos: Fundación Atapuerca, Los cazadores recolectores del Pleistoceno y del Holoceno en Iberia y el estrecho de Gibraltar: estado actual del conocimiento del registro arqueológico (J. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro, J.L. Arsuaga, Eds.), Burgos, 2014, pp. 485-488.
BELMAKER, M. (2010). On the road to China: The environmental landscape of the Early Pleistocene in western Eurasia and its implication for the dispersal of Homo. Vertebrate Paleobiology and Paleoanthropology Series, Ed. Springer, Asian Paleoanthropology: From Africa to China and Beyond (C.J. Norton, D.R. Braun, J.W.K. Harris, Eds.), 2010, pp. 31-40.
BOKELMANN, L., HAJDINJAK, M., PEYRÉGNE, S., BRACE, S., ESSEL, E., DE FILIPPO, C., GLOCKE, I., GROTE, S., MAFESSONI, F., NAGEL, S., KELSO, J., PRÜFER, K., VERNOT, B., BARNES, I., PÄÄBO, S., MEYER, M., STRINGER, C. (2019). A genetic analysis of the Gibraltar Neanderthals. PNAS, 116:31, 2019, pp. 1-6.
BORJA, C., GARCÍA-PACHECO, M., OLIVARES, E.G., SCHEUENSTUHL, G., LOWENSTEIN, J.M. (1997). Inmunospecifity of Albumin Detected in 1.6 Million-Year-Old fossils from Venta Micena in Orce, Granada, Spain. American Journal of Physical Anthropology, 103, 1997, pp. 433-441.
CARBONELL, V. (1965). The teeth of the neanderthal child from Gibraltar: A re-evaluation. Am. J. Phys. Anthrop., 23, 1965, pp. 41-50.
CARRIÓN, J.S., MUNUERA, M., NAVARRO, C. (1998). The palaeoenvironment of Carihuela Cave (Granada, Spain): a reconstruction on the basis of palynological investigations of cave sediments. Review of Palaeobotany and Palynology, 99, 1998, pp. 317-340.
CARRIÓN, J.S., FERNÁNDEZ, S., JIMÉNEZ-ARENAS, J.M., MUNUERA, M., OCHANDO, J., AMORÓS, G., PONCE DE LEÓN, M., ZOLLIKOFER, C., MARTÍN-LERMA, I., TORO-MOYANO, I., HAJDAS, I., WALKER, M.J. (2019). The sequence at Carihuela Cave and its potential for research into neanderthal ecology and the Mousterian in southern Spain. Quaternary Science Reviews, 217, 2019, pp. 194-216.
COLE, J. (2017). Assessing the calorific significance of episodes of human cannibalism in the Palaeolithic. Scientific Reports, 7:44707, 2017, pp. 1-10.
DENNELL, R., ROEBROEKS, W. (2005). An Asian perspective on Early Human Dispersal from Africa. Nature, 438, 2005, pp. 1099-1104.
DE LUMLEY, M.A. (1973). Anténéandertaliens et Néandertaliens du bassin mèditerranéen occidental européen. Études Quaternaires, 2, Editions du Laboratoire de Paleontologie Humaine et de Prehistoire. Université de Provence, 1973, pp. 247-309.
FIORENZA, L., BENAZZI, S., ESTALRRICH, A., KULLMER, O. (2020). Chapter 3: Diet and cultural diversity in Neanderthals and modern humans from dental macrowear analyses. Academic Press, Dental wear in evolutionary and biocultural contexts (C.W. Schmidt, J.T. Watson, Eds.), 2020, pp. 39-72.
GARCÍA-SÁNCHEZ, M. (1960). Restos humanos del paleolítico medio y superior y del neo-eneolítico de Piñar (Granada). Trabajos del Instituto <Bernardino de Sahagún> de Antropología y Etnología, 15:2, 1960, pp. 17-78.
GARCÍA-SÁNCHEZ, M., TILLIER, A.M., GARRALDA, M.D., VEGA-TOSCANO, G. (1994). Les dents d’enfant des niveaux moustériens de la grotte de Carihuela (Grenade-Espagne). Paléo, 6, 1994, pp. 79-88.
GARRALDA, M.D. (1997). The human paleontology of the Middle to Upper Paleolithic Transition on the iberia peninsula. Nueva York, Ed. Aldine de Gruyter, Conceptual issues in modern human origins research (G.A. Clark, C.M. Willermet, Eds.), 1997, pp. 148-160.
GARRALDA, M.D. (2005). Los neandertales en la península ibérica. Munibe, 57, 2005, pp. 290-314.
GARROD, D.A.E; DUDLEY-BUXTON, L.H; ELLIOT-SMITH, G; BATE, D.M.A; SPILLER, R.C; HINTON, M.A.C. y FISCHER, P. (1928). Excavation of a Mousterian Rock-Shelter at Devil’s Tower, Gibraltar. The Journal of the Royal Anthropological Institute of Great Britain and Ireland, 58, 1928, pp. 33-113.
GIBERT, J., PALMQVIST, P., (1995). Fractal analysis of the Orce skull sutures. Journal of Human Evolution, 28, 1995, pp. 561-575.
GIBERT, J., GIBERT, Ll., ALBADALEJO, S., RIBOT, F., SÁNCHEZ, F., GIBERT, P. (1999). Molar tooth fragment BL5-0: the oldest human remain found in the Plio-Pleistocene of Orce (Granada Province, Spain). Human Evolution, 14:1-2, 1999, pp. 3-19.
GILES-GUZMÁN, F., LEE-HOLMES, T. (2018). The Vanguard cave child. Third neanderthal fossil discovered in Gibraltar’s new world heritage site. Ice Age Europe, Network of heritage sites magazine (K. Hieke, G.C. Weniger, N. Powe, Coords.), 2018, pp. 12-14.
GODINHO, R.M., FITTON, L.C., TORO-IBACACHE, V., STRINGER, C.B., LACRUZ, R.S., BROMAGE, T.G., O’HIGGINS, P. (2018). The biting perfomance of Homo sapiens and Homo heidelbergensis. Journal of Human Evolution, 118, 2018, pp. 56-71.
HABER-URIARTE, M. (2002). Fragmento de hueso frontal del musteriense de Cueva Horá (Granada, España). Zephyrus, 55, 2002, pp. 323-329.
JIMÉNEZ-ARENAS, J.M., MARTÍN-LAGOS, I. (2014). Cueva Horá (Darro, Granada): Pleistoceno Inferior y Medio de la Cuenca de Guadix-Baza y Orce. Universidad de Burgos: Fundación Atapuerca, Los cazadores recolectores del Pleistoceno y del Holoceno en Iberia y el estrecho de Gibraltar: estado actual del conocimiento del registro arqueológico (J. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro, J.L. Arsuaga, Coords.), Burgos, 2014, pp. 493-494.
JIMÉNEZ-ARENAS, J.M., PALMQVIST, P., ESPIGARES, Mº P., ROS-MONTOYA, S., MARTÍNEZ-NAVARRO, B. (2016). La especie Dispersa(da). ¿Quiénes somos? ¿De dónde venimos? Junta de Andalucía, Orce: Homininos, hienas, mamuts y otras bestias (B. Martínez Navarro, R. Sala Ramos, Coords.), Granada, 2016, pp. 245-274.
JIMÉNEZ-ARENAS, J.M., BIENVENU, T., TORO-MOYANO, I., PONCE DE LEÓN, M., ZOLLIKOFER, C. (2019). Virtual reconstruction and re-evaluation of the Neanderthal frontal bone from Carigüela Cave (Granada, Spain). Quaternary Science Reviews, 217, 2019, pp. 89-97.
LACRUZ, R.S., BROMAGE, T.G., O’HIGGINS, P., ARSUAGA, J.L., STRINGER, C., GODINHO, R.M., WARSHAW, J., MARTÍNEZ, I., GRACIA-TELLEZ, A., BERMÚDEZ DE CASTRO, J.M., CARBONELL, E. (2015). Ontogeny of the maxilla in neanderthals and their ancestors. NATURE Communications, 6:8996, 2015, pp. 1-6.
MARTÍNEZ-NAVARRO, B. (2002). The skull of Orce: parietal bones or frontal bones? Journal of Human Evolution, 42, 2002, pp. 265-270.
MICHEL, V., SHEN, G., SHEN; C-C., FORNARI, M., VÉRATI, C., GALLET, S., SABATIER, D. (2011). Les derniers Homo heidelbergensis et leurs descendants les néandertaliens: datation des sites d’Orgnac 3, du Lazaret et de Zafarraya. Comptes Rendus Palevol, 10, 2011, pp. 577-587.
MONCLOVA-BOHÓRQUEZ, A., BARROSO-RUIZ, C., CAPARRÓS, M., MOIGNE, A.M. (2012). Una aproximación a la comprensión de la fauna de macromamíferos de la cueva de Zafarraya (Alcaucín, Málaga). Menga, Evolución humana y poblamiento del Pleistoceno en Andalucía (J.M. Jiménez Arenas, Ed.), 3, Granada, 2012, pp. 83-106.
MONTES-GUTIÉRREZ, R. (2013). Paleolítico Inferior Arcaico en la península ibérica. Tiempo y Sociedad, 13, 2013, pp. 5-44.
NABAIS, M., ZILHAO, J. (2019). The consumption of tortoise among Last Interglacial Iberian Neanderthals. Quaternary Science Reviews, 217, 2019, pp. 225-246.
PALMQVIST, P. (1997). A critical re-evaluation of the evidence for the presence of hominids in Lower Pleistocene times at Venta Micena, Southern Spain. Journal of Human Evolution, 33, 1997, pp. 83-89.
PANIAGUA-PÉREZ, J.P. (1998). El final de los complejos musterienses y los comienzos del Paleolítico Superior en el sur de la península ibérica. SPAL, 7, 1998, pp. 45-67.
RAMOS-FERNÁNDEZ, J. (1982). Los materiales del nivel Musteriense de la Cueva de Belda (Cuevas de San Marcos, Málaga). Zephyrus, 34-35, 1982, pp. 17-25.
RAMOS-MUÑOZ, J. (1998). La ocupación prehistórica de los medios kársticos de montaña en Andalucía. Madrid, Instituto Tecnológico Geominero de España (J.J Durán, J. López Martínez, Eds.), Karst en Andalucía, 1998, pp. 63-84.
RIBOT, F., GIBERT, L., FERRÀNDEZ-CAÑADELL, C., GARCÍA-OLIVARES, E., SÁNCHEZ, F., LERÍA, M. (2015). Two deciduous human molars from the Early Pleistocene deposits of Barranco León (Orce, Spain). Current Anthropology, 56:1, 2015, pp. 134-142.
RIVERA-ARRIZABALAGA, A. (2008a). Relación entre neandertales y cromañones: un enfoque cognitivo. Zephyrus, 61, 2008a, pp. 85-106.
RIVERA-ARRIZABALAGA, A. (2008b). Cognición y conducta de neandertales y humanos modernos. Revista portuguesa de Arqueologia, 11:1, 2008b, pp. 5-32.
RODRÍGUEZ-CUENCA, J.V. (2003). Cap. VIII – Presión ambiental, dieta y enfermedades dentales. Bogotá, Universidad Nacional de Colombia, (J.V. Rodríguez Cuenca, Coord.), Dientes y diversidad humana: avances de la antropología dental, 2003, pp. 117-138.
SALA-RAMOS, R., TORO-MOYANO, I., BARSKY, D., MENÉDEZ-GRANDA, L., MORILLA-MENESES, A., TORRENTE-CASADO, R., PINTO-ANACLETO, A., CHACÓN-NAVARRO, G., GÓMEZ-MERINO, G., CAUCHE, D., CELIBERTI, V., GRÉGOIRE, S., MONCEL, M.H., DE LUMLEY, H., LEBÈGUE, F., AGUSTÍ-BALLESTER, J., JIMÉNEZ-ARENAS, J.M., MARTÍNEZ-NAVARRO, B., OMS-LLOBET, O., TARRIÑO-VINAGRE, A. (2011). Orígenes de la ocupación humana de Europa: Guadix-Baza y Orce. Menga, 02, 2011, pp. 111-133.
SALA-RAMOS, R., MENÉNDEZ-GRANDA, L., ROS-MONTOYA, S., TORO-MOYANO, I. (2014). Barranco León: Pleistoceno Inferior y Medio de la Cuenca de Guadix-Baza y Orce. Universidad de Burgos: Fundación Atapuerca. Los cazadores recolectores del Pleistoceno y del Holoceno en Iberia y el estrecho de Gibraltar: estado actual del conocimiento del registro arqueológico (J. Sala Ramos, E. Carbonell, J.M. Bermúdez de Castro, J.L. Arsuaga), Burgos, 2014, pp. 477-478.
SALA-RAMOS, R., BARSKY, D., MENÉNDEZ-GRANDA, L., VERGÈS, J.M., TITTON, S., TARRIÑO, A., CÁNOVAS, I., SÁNCHEZ, L. (2016). ¿Qué herramientas utilizaba el hombre de Orce? Las industrias líticas arcaicas de los yacimientos de Barranco León y Fuente Nueva 3 en el contexto de la primera ocupación de Europa. Orce: Homininos, hienas, mamuts y otras bestias (B. Martínez Navarro, R. Sala Ramos, Eds.), Junta de Andalucía, Granada, 2016, pp. 217-244.
SÁNCHEZ, F. (1999). Comparative biometrical study of the Mousterian mandible from Cueva del Boquete de Zafarraya (Málaga, Spain). Human Evolution, 14:1-2, 1999, pp. 125-138.
STRINGER, C.B., FINLAYSON, J.C., BARTON, R.N.E., FERNÁNDEZ-JALVO, Y., CÁCERES, I., SABIN, R.C., RHODES, E.J., CURRANT, A.P., RODRÍGUEZ-VIDAL, J., GILES-PACHECO, F., RIQUELME-CANTAL, J.A. (2008). Neanderthal exploitation of marine mammals in Gibraltar. PNAS, 105:38, 2008, pp. 14.319-14.324.
TITTON, S., BARSKY, D., BARGALLÓ, A., SERRANO-RAMOS, A., VERGÈS, J.P., TORO-MOYANO, I., SALA-RAMOS, R., GARCÍA-SOLANO, J., JIMÉNEZ-ARENAS, J.M. (2020). Subspheroids in the lithic assemblage of Barranco León (Spain): Recognizing the late Oldowan in Europe. PLoS ONE, 15:1, 2020, pp. 1-41.
TORO-IBACACHE, V., MUÑOZ, V.Z., O’HIGGINS, P. (2015). The relationship between skull morphology, masticatory muscle force and cranial skeletal deformation during biting. Annals of Anatomy, 2015, pp. 1-26.
TORO-MOYANO, I., BARSKY, D., CAUCHE, D., CELIBERTI, V., GRÈGOIRE, S., LEBEGUE, F., MONCEL, M.H., DE LUMLEY, H. (2011). The archaic stone tool industry from Barranco León and Fuente Nueva 3, (Orce, Spain): Evidence of the earliest hominin presence in southern Europe. Quaternary International, 243, 2011, pp. 80-91.
TORO-MOYANO, I., MARTÍNEZ-NAVARRO, B., AGUSTÍ, J., SOUDAY, C., BERMÚDEZ DE CASTRO, J.M., MARTINÓN-TORRES, M., FAJARDO, B., DUVAL, M., FALGUÈRES, C., OMS, O., PARÉS, J.M., ANADÓN, P., JULIÀ, R., GARCÍA-AGUILAR, J.M., MOIGNE, A-M., ESPIGARES, M.P., ROS-MONTOYA, S., PALMQVIST, P. (2013). The oldest human fossil in Europe, from Orce (Spain). Journal of Human Evolution, 65, 2013, pp. 1-9.
URQUIZA, R., BOTELLA, M., CIGES, M. (2005). Study of a temporal bone of Homo Heidelbergensis. Acta Oto-Laryngologica, 125, 2005, pp. 457-463.
VAESEN, K., SCHERJON, F., HEMERIK, L., VERPOORTE, A. (2019). Inbreeding, Allee efects and stochasticity might be sufficient to account for Neanderthal extinction. PLoS ONE, 14:11, 2019, pp. 1-15.
VILLALOBOS-MEGÍA, M., BRAGA-ALARCÓN, J.C. (2003). Capítulo 9 – Cuenca de Guadix-Baza. Málaga, Junta de Andalucía, 2003, pp. 199-213.
VILLAR-CALVO, A. (1998). La pervivencia del Musteriense en la mitad sur peninsular durante el Pleniglacial Superior. Una revisión crítica de las dataciones. Espacio, Tiempo y Forma, Serie I, Prehistoria y Arqueología, 11, 1998, pp. 57-64.
WOOD, R.E., BARROSO-RUIZ, C., CAPARRÓS, M., JORDÁ-PARDO, J.F., GALVÁN-SANTOS, B., HIGHAM, T.F.G. (2013). Radiocarbon dating casts doubt on the late chronology of the Middle to Upper Palaeolithic transition in southern Iberia. PNAS, 110:8, 2013, pp. 2781-2786.
ZILHAO, J., ANGELUCCI, D.E., ARAÚJO-IGREJA, M., ARNOLD, L.J., BADAL, E., CALLAPEZ, P., CARDOSO, J.L., D’ERRICO, F., DAURA, J., DEMURO, M., DESCHAMPS, M., DUPONT, C., GABRIEL, S., HOFFMANN, D.L., LEGOINHA, P., MATIAS, H., MONGE-SOARES, A.M., NABAIS, M., PORTELA, P., QUEFFELEC, A., RODRIGUES, F., SOUTO, P. (2020). Last interglacial Iberian neandertals as fisher-hunter-gatherers. Science, 367, 2020, pp. 1443-1457.
