Jim Marshall
Department of Earth Sciences University of Liverpool, 4, Brownlow St, Liverpool L69 3GP.
Email - isotopes@liv.ac.uk.
Almost all biotic events throughout the geological record are accompanied by stratigraphic changes in carbon and oxygen stable isotope signatures that are recorded in contemporaneous marine sediments and fossils. Isotopic records from bulk sediments often reflect the overall pattern of changes in the marine environment – but the magnitude of the changes is commonly masked by the effects of diagenesis. Quantitiative interpretations should be based, wherever possible, on analyses of prisitine fossil material (Marshall, 1992).
In the best-preserved calcareous and phosphatic fossils but almost never in whole rock samples, oxygen isotope (d18O) data can identify patterns of change in temperature and ocean water composition (ice-volume and, more locally, salinity). Temperature change has been shown to be a significant contributor to biotic events at the End Ordovician and late Devonian (Brenchley et al., 1994,1995; Joachimski and Buggisch, 2002).
Carbon isotope values (d13C) from rocks, fossils and biomarkers record changes in the carbon cycle. Often signals can be shown to reflect global changes (Marshall et al., 1997) but local amplification effects must be considered. Carbon–cycle changes include changes in carbon inputs (eg from rivers) or outputs (especially methane), organic productivity, and carbon burial (Holser, 1997). Attempts to utilise carbon isotope values in associated carbonates and organics has enomous potential but the proxies may not work in very high levels of pCO2 inferred for much of the past (Joanchimski et al., 2002). All five of the major Phanerozoic mass extinction events are matched by significant carbon isotopic excursions. Positive excursions such as those associated with the end Ordovician, and late Devonian are generally attributed to increases in organic carbon burial or productivity - perhaps associated with enhanced nutrient fluxes. Negative excursions found associated with the end Permian and the Palaeocene/Eocene boundary are alternatively interpreted to record collapse in primary productivity or massive release of methane from hydrates (Holser, 1997; Thomas et al., 2002). Detailed comparison of isotopic signatures from benthic and planktonic faunal elements at the K/T boundary enable changes in ocean structure to be monitored (eg. Kaiho et al., 1999). Carbon isotope values can be affected by many environmental factors so although models, including box models and more sophisticated circulation and energy balance models, can be used to help constrain the interpretations it is often difficult to quantify the changes in the past environments with certainty.
Detailed, high-resolution, stratigraphic comparisons of faunal and isotopic data are particularly valuable in helping to resolve some of the relationships between bioevents and palaeoceanography. Isotopic values can provide a geologically instantaneous chronological marker (cf. Thomas et al., 2002) of global environmental change that may be recorded in sediments with very different faunas in different areas of the world (Brenchley et al., 2003). This can enable detailed correlation between disparate sites at a resolution that exceeds that of biostratigraphy. The detailed sequence of changes within a biotic event can be determined (Fig 1).
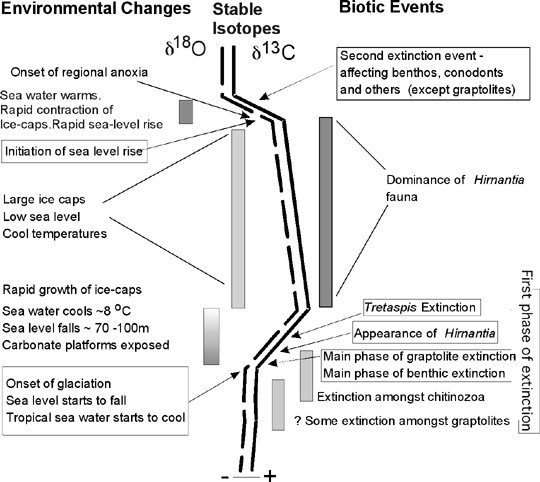
Figure 1. End Ordovician Extinctions and related environmental changes. Recent work (eg. Marshall et al., 1997, Brenchley et al., 2003) has shown that the stable isotopic record from sediments and fossils can provide a global chronostratigraphic ruler that enables global correlation of biotic events and environmental change. The diagram shows an idealised carbon and oxygen stable isotopic profile based on data from Baltica and Laurentia. The positive excursions in both d18O and d13C are synchronous and correspond to changes in temperature and carbon cycling associated with the growth and demise of the Gondwanan ice caps. The biotic changes, which together comprise the end-Ordovician mass-extinction are concentrated in two main phases associated with the most rapid enmvrironmental changes. The isotopic data provide key evidence for the magnitude of environmental change and the relationship with faunal occurrences enables the detailed 'architecture' of the biotic changes to be determined. Despite the massive scale of the environmental disruption at the end of the Ordovician the degree of ecological disruption was relatively small (Brenchley et al., 2001). (Figure is based on Brenchley et al., 2003, Fig. 15).
Isotopic records be used to distinguish between mass extinction events where environmental changes accompany, and potentially drive, biotic change (such as the end-Ordovician) from those events where the isotopic values, environmental and faunal changes are responses to an external driving mechanism, such as a bolide impact.
References
Brenchley, P. J., Carden, G.A., Hints, L., Kaljo, D., Marshall, J. D., Martma,T., Meidla, T. and Nõlvak, J. 2003. High-resolution isotope stratigraphy of Late Ordovician sequences: constraints on the timing of bio-events and environmental changes associated with mass extinction and glaciation. Geological Society of America Bulletin, 115, 89-104.
Brenchley, P.J, Carden, G.A.F. and Marshall, J.D. 1995. Environmental changes associated with the "first strike" of the late Ordovician mass extinction. Modern Geology, 20, 69-82.
Brenchley, P.J., Marshall, J.D. and Underwood, C.J. 2001. Do all mass extinctions represent an ecological crisis? Evidence from the late Ordovician. Geological Journal, 36, 329-340.
Brenchley, P.J., Marshall, J.D., Robertson, D.B.R. Carden, G.A.F., Long, D.G.F., Meidla, T., Hints, L. and Anderson, T.F. 1994. Bathymetric and isotopic evidence for a short-lived late Ordovician glaciation in a greenhouse period. Geology, 22, 295-298.
Joachimski, M.M. and Buggisch, W. 2002. Conodont apatite delta O-18 signatures indicate climatic cooling as a trigger of the Late Devonian mass extinction. Geology, 30, 711-714.
Joachimski, M.M., Pancost, R.D., Freeman, K.H., Ostertag-Henning, C., Buggisch, W. 2002. Carbon isotope geochemistry of the Frasnian-Famennian transition Palaeogeography Palaeoclimatology Palaeoecology, 181, 91-109.
Kaiho, K., Kajiwara, Y., Tazaki, K., Ueshima, M., Takeda, N., Kawahata, H., Arinobu, T., Ishiwatari, R., Hirai, A. and Lamolda, M.A. 1999. Oceanic primary productivity and dissolved oxygen levels at the Cretaceous/Tertiary boundary: Their decrease, subsequent warming, and recovery. Paleoceanography, 14, 511-524.
Holser, W.T 1997.Geochemical events documented in inorganic carbon isotopes. Palaeogeography Palaeoclimatology Palaeoecology, 132, 173-182.
Marshall, J.D., Brenchley, P.J., Mason, P., Wolff, G.A., Astini, R.A., Hints, L., Meidla, T. 1997. Global carbon isotopic events associated with mass extinction and glaciation in the Late Ordovician. Palaeogeography, Palaeoclimatology, Palaeoecology, 132, 195-210.
Marshall, J.D. 1992. Climatic and oceanographic isotopic signals from the carbonate rock record and their preservation. Geological Magazine, 129,143-160.
Thomas, D.J., Zachos, J.C., Bralower, T.J., Thomas, E. and Bohaty, S. 2002.Warming the fuel for the fire: Evidence for the thermal dissociation of methane hydrate during the Paleocene-Eocene thermal maximum. Geology, 30, 1067-1070.