
ANÁLISIS TAFONÓMICO DE LOS RESTOS FAUNÍSTICOS DEL COMPLEJO ESTRUCTURAL XV DEL POLIDEPORTIVO DE MARTOS, JAÉN
TAPHONOMIC ANALYSIS OF THE FAUNAL REMAINS OF THE STRUCTURAL COMPLEX XV OF ‘POLIDEPORTIVO DE MARTOS’, JAEN
Álvaro LÓPEZ DE LOS MOZOS MOSQUERA*
Resumen
En este trabajo se presentan los resultados del exhaustivo análisis zooarqueológico y tafonómico del conjunto faunístico del Complejo Estructural XV del Polideportivo de Martos, Jaén. A partir de los resultados obtenidos, el objetivo del estudio tratará de lograr un acercamiento a los conocimientos socio-económicos y patrones de explotación y consumo animal de estas sociedades en el Neolítico Reciente en la Alta Andalucía.
Palabras clave:
Zooarqueología, Tafonomía, Neolítico Reciente, Alta Andalucía, Martos
Abstract
This paper presents the results of the exhaustive zooarchaeological and tafonomic analysis of the faunal complex of the Structural Complex XV of the ‘Polideportivo de Martos’, Jaen. Based on the results obtained, the objective of the study will be to propose a first approach to the socio-economic knowledge and patterns of exploitation and animal consumption of these societies in the Recent Neolithic in Upper Andalusia.
Key words:
Zooarchaeology, Taphonomy, Recent Neolithic, Upper Andalusia, Martos
INTRODUCCIÓN
El conjunto arqueológico, prehistórico, del Polideportivo de Martos, inscrito en el Catálogo General del Patrimonio Histórico de Andalucía como Bien de Interés Cultural (BIC) (Riquelme et al., 2012), se sitúa en la localidad de Martos (Jaén), en las coordenadas UTM 414635,4174490 (AFONSO et al., 2014) y con una extensión de 57,1963 ha (RIQUELME et al., 2012) (Fig. 1).
Fig. 1. Ubicación de Martos, Jaén
El yacimiento se localiza dentro de la Campiña Alta Occidental, la cual se sitúa entre las cotas de 400 m. y 600 m. sobre una gran loma con una inclinación suave en dirección Sur-Norte hacia el Arroyo de la Fuente, formando una pequeña vega en su curso medio (LIZCANO et al., 1991).
El yacimiento se caracteriza por contener estructuras subterráneas de diversas configuraciones y dimensiones (AFONSO et al., 2014).
En su gran mayoría las estructuras que abundan, además de los tramos de foso, son fosas piriformes, conservadas exclusivamente en su parte inferior y a menudo superpuestas, cuyo relleno estratigráfico es el resultado de distintos usos de espacio. Así pues, tanto la cercanía de las estructuras como el material hallado y recuperado, nos informa que la unidad residencial, que debió existir, contó con complejos articulados entre sí (AFONSO et al. 2014).
Para el estudio del yacimiento del Polideportivo de Martos, se realizó una matriz estructural, teniendo en cuenta la estratigrafía horizontal, donde se pudieron definir tres grandes fases estructurales.
La cronología del yacimiento, se determinó por la toma de quince muestras, de las cuales trece correspondían a restos óseos faunísticos, una muestra perteneciente a un carbón de hogar y un último resto humano (AFONSO et al. 2014).
Las propuestas cronológicas para el conjunto de la ocupación investigada en el yacimiento nos situarían en los siguientes intervalos: 3514-2350 cal ANE (Antes de Nuestra Era) por Calib 7.0.2; 3520-2467 cal ANE por combinación mediante el método del error mínimo global; 2299-3462 cal ANE por Bcal; y 3472-2079 cal ANE por OxCal, sugiriéndonos que la fase IA se desarrolló a principios de la segunda mitad del IV milenio ANE, la fase IB a principios del último cuarto del IV milenio ANE y la fase IIA en el último tercio del IV milenio ANE (AFONSO et al. 2014).
El CE XV, objeto de estudio del presente trabajo, es una de las áreas estructurales más importantes del Polideportivo de Martos, perteneciente a las Fases I y II.
La actividad antrópica en el Complejo Estructural XV, está constituida por las Fases IA, IB, IIA y IIB (Fig. 2).
• Fase Ia. Sobre el suelo, se encuentra el primer nivel donde se halló un esqueleto en conexión anatómica de un perro, sobre una capa de yeso de 3 centímetros de grosor, cubriendo exclusivamente el cuadrante noreste del fondo (UE 7a).
Sobre este primer nivel de “fundación” se halla una capa compuesta por materiales de desecho (UE 7) con recipientes cerámicos como ollas globulares, cazuelas, vasos, entre otros, así como abundantes restos de fauna.
• Fase Ib. Superpuesta a la capa anterior, aparece una nivelación del espacio con margas apisonadas (UE 6a), sirviendo como nuevo suelo con características muy parecidas al anterior pero con escasa presencia de materiales cerámicos.
• Fase IIa. En esta subfase se construye un banco de piedra corrido en todo el espacio de la estructura, constituido por margas apisonadas y piedrecillas embutidas para dar una mejor resistencia (UE 5).
Sobre este banco se halla un relleno por acumulación de restos faunísticos y útiles líticos.
• Fase IIb. En esta subfase se realiza una nueva nivelación (UE 4) con una recubrimiento de tierra apisonada de unos 4 centímetros de grosor, creando así una nueva plataforma cubriendo el anterior banco, el cual pierde su función.
Es en este nivel donde aparece el esqueleto completo de un ternero, de seis meses, junto a una acumulación de ceniza, relacionadas con un hogar. Abundantes son también, en este nivel, los restos de fauna y materiales líticos.
Tras esta capa, la ocupación antrópica desaparece colmatada por un depósito de abandono (UE 3A), con gran cantidad de piedras de reducido tamaño, así como cerámicas fragmentadas y trozos de molino (LIZCANO et al. 1991).
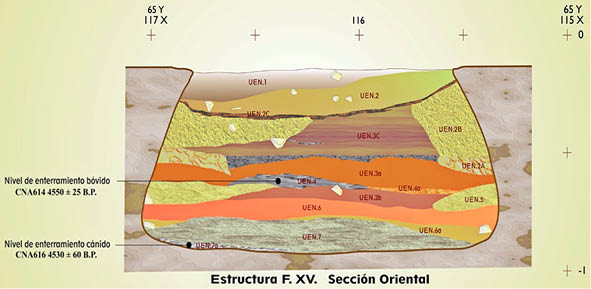
Fig. 2. Sección del Complejo Estructural XV (Afonso et al., 2014).
METODOLOGÍA DE ESTUDIO
Los materiales óseos analizados para el presente trabajo corresponden a la campaña de excavación del Complejo Estructural XV del Polideportivo de Martos 1991. Dichos materiales han sido separados por su correspondiente fase (Fase I o Fase II) con un número total de 1.173 restos óseos analizados.
El proceso de identificación anatómica y taxonómica, así como el estudio tafonómico se realizó con la ayuda de diversos Atlas osteológicos y tafonómicos (FERNÁNDEZ-JALVO Y ANDREWS 2016) y colaboración con el Dr. J.A. Riquelme Cantal, así como el empleo de un microscopio USB de x1600 aumentos, una báscula de precisión y un calibre pie de rey.
Para ello, inicialmente se llevó a cabo un proceso de identificación anatómica y taxonómica, para el revisado y la contrastación de los datos publicados con anterioridad (LIZCANO et al. 1991; RIQUELME et al. 2012), habiendo podido incluir dos nuevos taxones nunca antes relacionados con el Conjunto Arqueológico del Polideportivo de Martos, y posteriormente se efectuó un análisis tafonómico para poder hallar cualquier alteración biótica (modificaciones humanas o animales) o abiótica (modificaciones climáticas, edáficas, diagenéticas, etc) en cada resto óseo.
Para un mayor análisis de cada elemento óseo se procedió a agrupar toda su información en los siguientes apartados: Nº, Nº de Siglado, Taxón, Parte Anatómica, Medidas (Largo, Ancho, Grosor), Peso, Observaciones.
Para los restos óseos indeterminables tanto a nivel taxonómico como anatómico, se procedió a una agrupación en huesos largos (HL) o huesos planos (HP), pertenecientes todos los restos a animales de talla media.
Por otra parte, los restos identificables fueron clasificados por especie. En el caso de la oveja y la cabra, todos los restos óseos que no se han podido determinar a nivel taxonómico, debido a su complejidad y similitud entre ambas especies, han sido incluidas en el grupo de los ovicaprinos (O/C).
Para una mejor comprensión de las gráficas y tablas se procedió a elaborar una serie de tablas con códigos representando al nombre científico de las especies, así como para el desglose anatómico (Tab. 1) (Tab. 2).
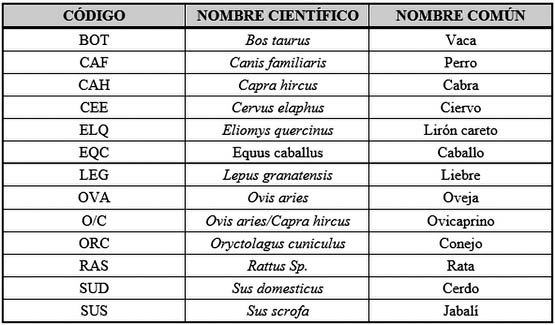
Tab. 1. Especies representadas en la muestra faunÍstica.
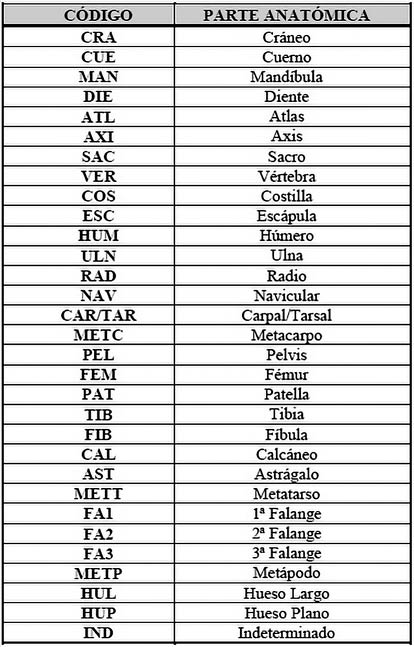
Tab. 2. Desglose anatómico representado en la muestra.
Una vez completada la identificación anatómica y taxonómica de la muestra faunística, se abordó la labor de la contabilización y ordenación de los restos analizados, usando los índices, más representativos, que permiten cuantificar la muestra ósea como: el Número de Restos (NR), el Número Mínimo de Elementos (NME), el Número Mínimo de Individuos (NMI).
Cada una de estas técnicas nos aportó una información detallada y complementaria con los demás índices, permitiéndonos una organización y un estudio exhaustivo del registro fósil analizado.
Para finalizar, en el análisis tafonómico se pudieron concretar diversas alteraciones bióticas (marcas de corte, alteraciones térmicas y marcas de carnívoros) y alteraciones abióticas como:
El weathering son alteraciones producidas por la exposición subaérea y son el resultado de la exposición del resto óseo a los agentes atmosféricos, como cambios de humedad, temperatura, exposición a los rayos ultravioleta, hasta que son enterrados, provocando exfoliaciones, agrietamientos y fisuras, pudiendo llegar a descomponer y desintegrar el hueso (BLASCO 2006; GARCIA SOLANO 2014).
Las oxidaciones son alteraciones producidas por acciones hídricas, donde el incremento de la humedad, así como, la presencia de aguas carbonatadas producen una serie de alteraciones que colorean la superficie del hueso con colores negruzcos pertenecientes a oxidaciones de óxidos de hierro y/o manganeso. (HEDGES Y MILARD 1995; GARCÍA SOLANO 2014)
Las vermiculaciones son unas alteraciones producidas por raíces que al entrar en contacto con la superficie del hueso provocan unas marcas en forma de ramificaciones, con sección en U, producidas por la acción de los ácidos carbónicos y cítricos presentes en las raíces de las plantas (GRAYSON 1988; FISHER 1995) (GIACOBONI Y PATOU-MATHIS 2002).
PRESENTACIÓN DE LOS DATOS
Análisis taxonómico y anatómico
Para llevar a cabo la cuantificación de la muestra se procedió a la agrupación del Número de Restos (NR) y el Número Mínimo de Individuos (NMI) por especies de macromamíferos y mesomamíferos de la Fase I y de la Fase II, por un lado, y de micromamíferos, por otro.
Se puede observar (Tab. 3) como la especie con mayor representación en la Fase I, en Número de Restos (NR), sería el perro (Canis familiaris) con 190 restos, todos ellos pertenecientes al enterramiento ritual de “fundación” de un solo individuo. Seguidamente encontraríamos a los ovicaprinos con 63 restos y 2 individuos; 14 restos de cabra (Capra hircus) y 1 individuo; 6 restos de oveja (Ovis aries) y 2 individuos; el cerdo tendría una representación de 18 restos y 2 individuos y la vaca (Bos taurus) con 2 restos y un Número Mínimo de Individuos (NMI) de 1 sería el animal doméstico con menor representación.
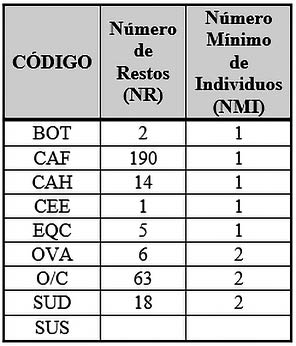
Tab. 3. Número de Restos Determinables (NRD) y Número Mínimo de Individuos (NMI) de macromamíferos y mesomamíferos de la Fase 1.
En cuanto a las especies salvajes, el número de restos es considerablemente menor al de los animales domésticos, como es el caso del ciervo (Cervus elaphus) con solo un resto determinable y por consiguiente un único individuo, así como el caballo (Equus caballus) con 5 restos determinables y un individuo.
El caballo entraría dentro del apartado de especies salvajes, dado que las hipótesis más aceptadas y estudios recientes (LIRA GARRIDO 2015; 2018) sugieren que la domesticación del caballo se produjo entre finales del II milenio A.N.E. y principios del I milenio A.N.E.
En la Fase II podemos observar como de los animales domésticos, los ovicaprinos son, con diferencia, la especie con mayor presencia en el conjunto faunístico, con un Número de Restos de 319 y 4 individuos entre los que se podría añadir un cabra (Capra hircus) con 6 restos y un individuo.
El resto de animales domésticos tiene una representación bastante menor, en comparación con los ovicaprinos, como es el caso del cerdo (Sus domesticus) con 31 restos de los cuales se han podido encontrar un Número Mínimo de 3 individuos; seguido del Canis familiaris, con 19 restos y un individuo.
La vaca (Bos taurus) se representa con 81 restos pero solo 6 restos determinables corresponden a un adulto consumido y los 75 restos restantes corresponden a una inhumación de un ternero de 6 meses.
Los animales salvajes están representados por el ciervo (Cervus elaphus) con 39 restos, en su mayoría fragmentos de cuerna, y un solo individuo; seguido por el caballo (Equus caballus) y el jabalí (Sus scrofa) con 3 restos y un individuo cada uno (Tab. 4).
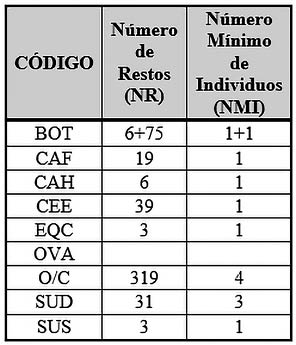
Tab. 4. Número de Restos Determinables (NRD) y Número Mínimo de Individuos (NMI) de macromamíferos y mesomamíferos de la Fase 2.
En cuanto a las especies incluidas en el apartado de microfauna, encontramos al conejo (Oryctolagus cuniculus) con una representación de 31 restos determinables y 2 individuos y a la liebre (Lepus granatensis) con 8 restos y un solo individuo, como posibles especies comestibles, debido a marcas de corte halladas; además de la rata (Rattus sp) con 4 restos y 2 individuos; al lirón con 2 restos determinables y un único individuo; y 35 restos óseos pertenecientes a un único individuo de roedor indeterminado hasta el momento (Tab. 5).
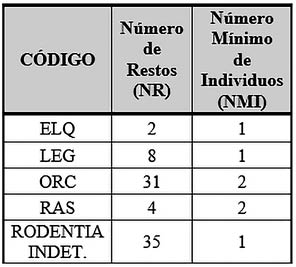
Tab. 5. Número de Restos Determinables (NRD) y Número Mínimo de Individuos (NMI) de la Microfauna Fase I y Fase II.
Dentro de la Fase I, las partes anatómicas con mayor representación (Tab. 6) (Fig. 3) serían las vértebras y costillas, que en su gran mayoría pertenecen al único individuo de Canis familiaris, seguido de las mandíbulas y dientes pertenecientes a dicho perro (Canis familiaris), a ovicaprinos y a cerdos (Sus domesticus).
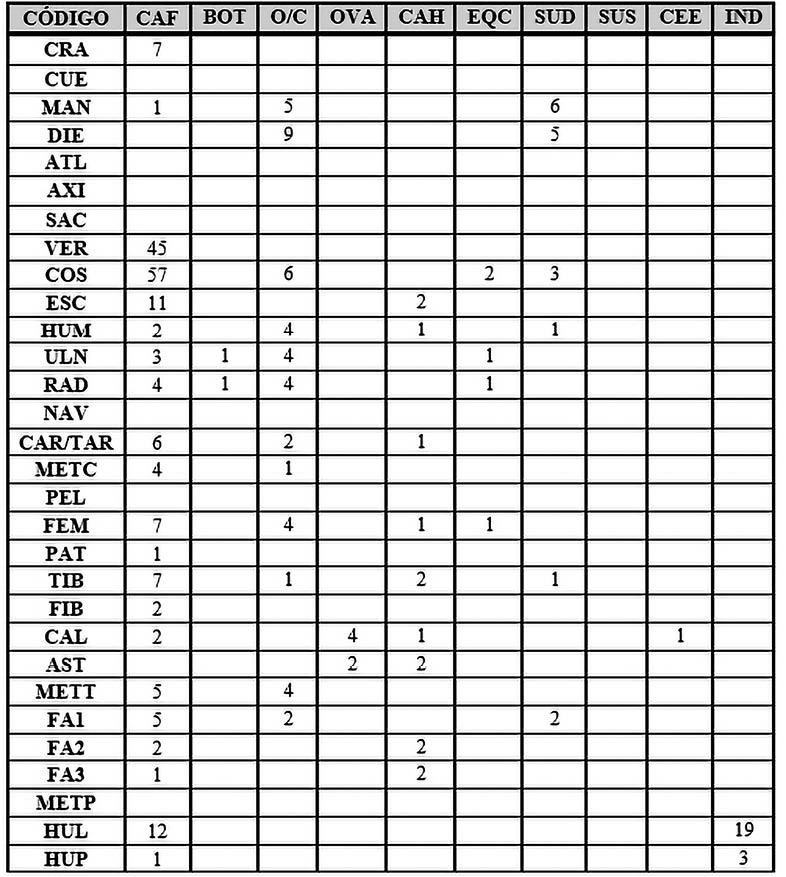
Tab. 6. Desglose anatómico de las especies de macromamíferos y mesomamíferos de la Fase 1.
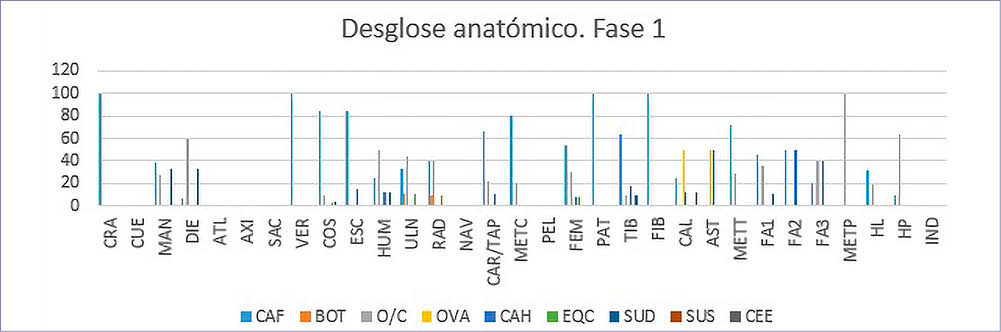
Fig. 3. Desglose anatómico NR% por especies de macromamíferos y mesomamíferos de la Fase 1.
Los elementos apendiculares están mayoritariamente representados por los ovicaprinos, lo que nos indicaría que las partes de cuartos traseros y delanteros era el principal alimento de esta sociedad, al menos así lo muestra el Complejo Estructural XV, indicándonos un fuerte control por la cabaña ovicaprina en estos momentos.
En cuanto a las partes anatómicas de los animales salvajes, solo han aparecido en esta fase 6 restos: 2 costillas, una ulna, un radio y un fémur pertenecerían al caballo (Equus caballus) y solo un calcáneo perteneciente a un ciervo (Cervus elaphus)
En cuanto a la Fase II (Tab. 7) (Fig. 4) la representación anatómica está protagonizada mayormente por los ovicaprinos (O/C) con 319 restos, de los cuales cabe destacar los elementos del esqueleto axial, como vértebras y costillas.
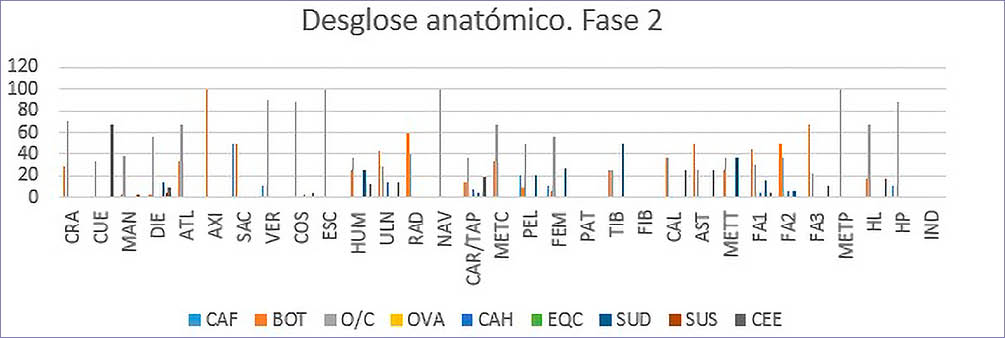
Fig. 4. Desglose anatómico NR% por especies de macromamíferos y mesomamíferos de la Fase 2.
En cuanto a la vaca (Bos taurus) cabe resaltar que los números están altamente influenciados por los restos del ternero inhumado en la Subfase IIB, apareciendo única y exclusivamente un fragmento de mandíbula, un diente, una costilla, un calcáneo, una falange proximal y un hueso largo pertenecientes a las partes anatómicas de una vaca consumida.
El cerdo (Sus domesticus) tomaría más presencia en esta fase en la dieta de esta sociedad, en comparación con los resultados de la Fase I.
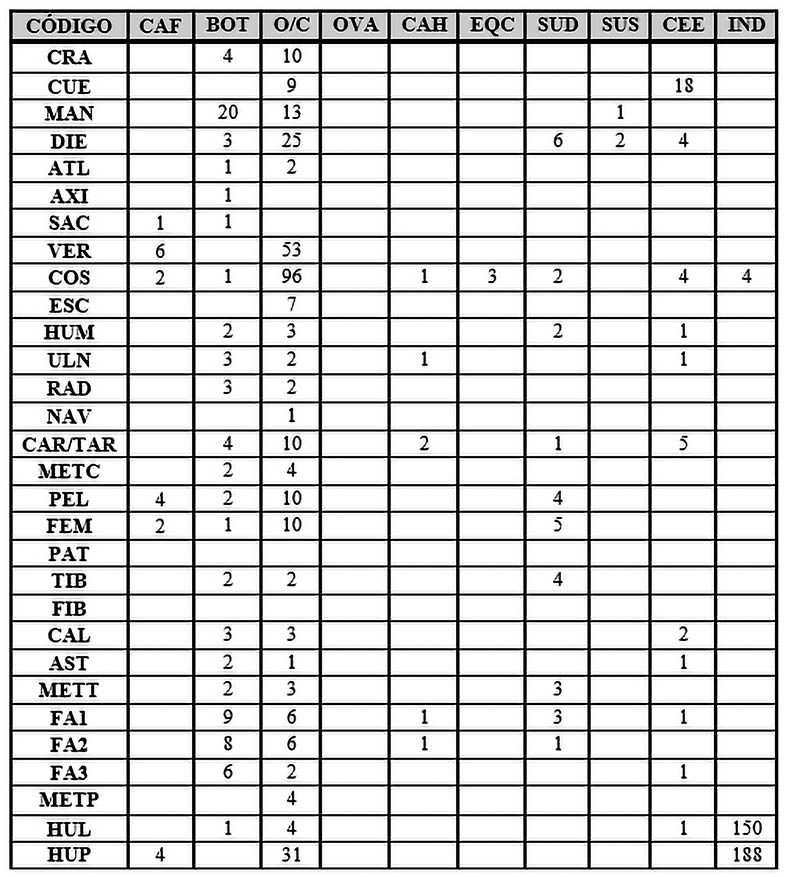
Tab. 7. Desglose anatómico de las especies de macromamíferos y mesomamíferos de la Fase 2.
En relación a los animales salvajes, es interesante destacar la presencia del jabalí (Sus scrofa) siendo posible su identificación gracias a dos restos dentales y un fragmento de mandíbula.
El ciervo también toma fuerza representativa con 39 restos, aunque a su vez este número tan elevado está influenciado por los 18 fragmentos de cuerna hallados. El resto de partes anatómicas del ciervo están conformadas por 4 dientes, 4 fragmentos de costillas, un húmero, una ulna, 5 huesos carpales/tarsales, 2 calcáneos, un astrágalo, una primera falange o falange proximal, una tercera falange o falange distal y un hueso largo sin poder ser identificado pero cuyas características son sin duda de un cérvido.
Por último, el caballo pierde importancia en comparación con los resultados de la Fase I, con solo 3 fragmentos de costillas.
Referente a las especies de micromamíferos que hemos podido analizar y afirmar su presencia, han podido ser determinados gracias a elementos anatómicos completos y particulares.
En el caso del conejo y la liebre las piezas óseas apendiculares tales como tibia, fémur y metápodos han servido para identificarlos como tales.
El resto de animales de pequeño tamaño, como la rata y el lirón han podido identificarse gracias a dos húmeros y a una mandíbula respectivamente. Estos animales, nunca antes habían podido ser relacionados con el Complejo del Polideportivo de Martos, Jaén.
Los 35 restos restantes representados en el apartado ROD, han podido ser identificados anatómicamente pero no taxonómicamente, es por ello que hemos decidido introducirlas en la sección de Rodentia (ROD) (Tab. 8).
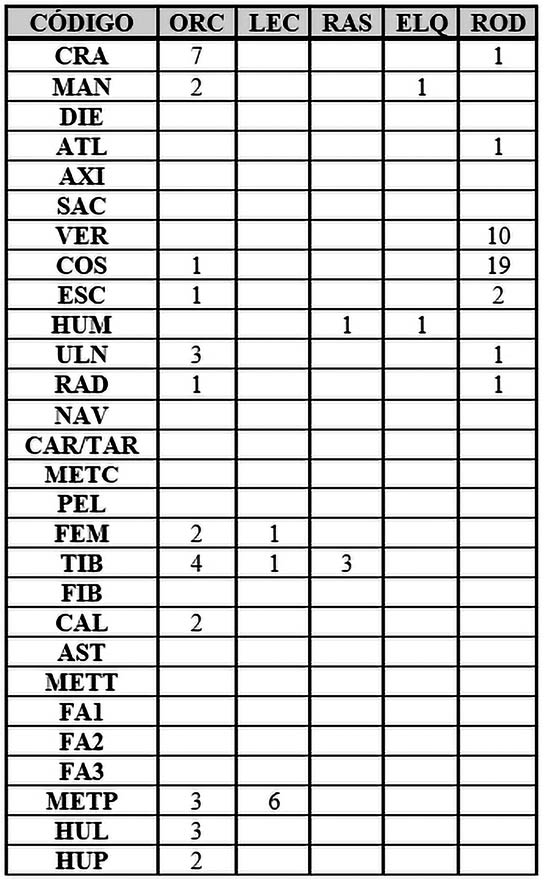
Tab. 8. Desglose anatómico de la fauna perteneciente a los micromamíferos de la Fase I y Fase II.
En cuanto a las edades de las especies identificadas, la gran mayoría entraría en la fase adulta, encontrando exclusivamente animales infantiles y juveniles en las especies de Bos taurus, los ovicaprinos y los cerdos (Sus domesticus).
En la fase I podemos encontrarnos con especímenes juveniles de las especies de ovicaprinos y cerdos.
En los ovicaprinos el individuo que ha podido ser identificado como juvenil, ha sido gracias a la no-fusión de los huesos apendiculares del fémur, húmero, así como una primera falange o falange proximal.
Respecto al cerdo (Sus domesticus), el único elemento que nos ha podido confirmar la presencia de un individuo juvenil ha sido la aparición de un premolar.
En la Fase II, siguen apareciendo animales subadultos tanto en ovicaprino y cerdo, así como el ternero hallado completo.
En los ovicaprinos, aparece un individuo juvenil, identificado por los huesos fragmentados de la escápula, calcáneo, dos metacarpos, dos fragmentos de fémur, un fragmento de mandíbula y una tibia. Así como un individuo infantil, reconocido por la presencia de una falange medial o segunda falange en el conjunto faunístico analizado.
En cuanto al cerdo, también aparece un individuo juvenil y otro infantil. Los huesos que han podido demostrar la presencia del individuo infantil son los fragmentos no fusionados de un fémur, una tibia, un húmero y elementos dentarios varios. El individuo infantil, ha podido ser identificado gracias a la presencia de un premolar deciduo.
Por último para la representación del peso de la fauna analizada y presumiblemente consumida se han descartado los dos enterramientos de animales en conexión anatómica, como son el caso del perro en la Fase I y el ternero en la Fase 2.
Los resultados obtenidos para la Fase I son los siguientes (Fig. 5):
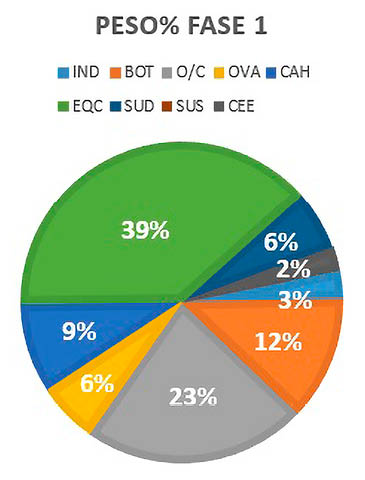
Fig. 5. Representación de Peso% por especies de macromamíferos y mesomamíferos de la Fase 1.
Los animales que más carne han podido aportar han sido el caballo con un 39% del peso de la muestra, seguido por los ovicaprinos y la vaca. Los restos indeterminados se sitúan en un 9% del peso analizado.
Los resultados obtenidos por la representación del Peso% en la Fase II sugieren que los ovicaprinos serían la principal fuente de alimento de los habitantes de este complejo con más del cincuenta por ciento de la dieta (entre O/C y CAH), seguido de los cerdos, indeterminados y cérvidos (Fig. 6).
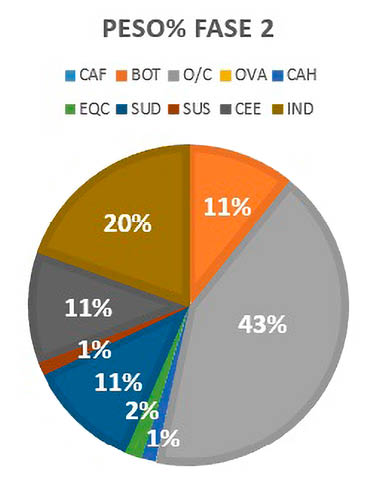
Fig. 6. Representación de Peso% por especies de macromamíferos y mesomamíferos de la Fase 2.
Análisis tafonómico
A continuación, se muestran una serie de tablas y gráficas donde se detallan las marcas y alteraciones bióticas y abióticas analizadas sobre el conjunto faunístico del Complejo Estructural XV, en las fases I y II (Tab. 9) (Tab. 10).
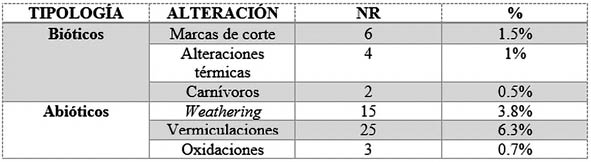
Tab. 9. Clasificación por alteraciones de Número de Restos (NR) y su % de la Fase 1.

Tab. 10. Clasificación por alteraciones de Número de Restos (NR) y su % de la Fase 2.
En cuanto a las alteraciones bióticas, las marcas de corte (BINFORD 1981; GARCÍA SOLANO 2014) constituyen el principal proceso tafonómico de acción antrópica sobre el conjunto analizado de la Fase I. En su caso, constituyen el 1.5% de todos los restos analizados de dicha fase, con un total de 6 restos con marcas de corte, y el 50% de las marcas bióticas analizadas.
Dentro de estas marcas, se han podido distinguir cuatro tipos, entre las que están las marcas de desollado, evisceración, desarticulación y descarnación.
Las marcas de descarnación junto a las de desarticulado, representan el 66% de las marcas de corte de la Fase I, determinadas en un húmero, un fémur y dos ulnas. El desollado y la evisceración constituyen en exclusiva los 34% restantes, distinguidas en un hueso plano y una costilla (Fig. 7).
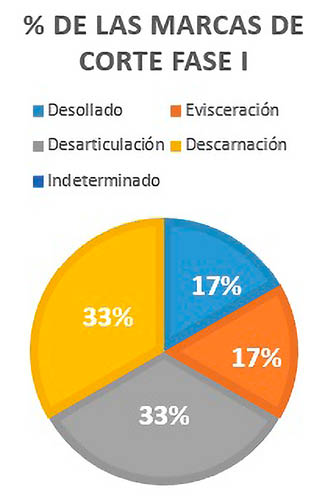
Fig. 7. Porcentajes de cada tipo de marca de corte analizada en la Fase I.
Las alteraciones térmicas, en la Fase I, suponen solo un 1% de todo el material analizado de dicha fase y el 33% de las marcas bióticas, con solo 4 restos quemados, con coloraciones grisáceas y blanquecinas en todo su conjunto, lo que podría indicarnos la intencionalidad de eliminación de estos residuos, y no una intención de cocinado.
Por último, las alteraciones producidas por carnívoros solo constituyen el 0.5% de todos los restos de la Fase I y el 17% de las marcas alteradas bióticamente, con solo dos marcas analizadas, posiblemente producidas por mordedura de perro (Canis familiaris) (SELVAGGIO 1994; DOMÍNGUEZ-RODRIGO Y PIQUERAS 2003; GARCÍA SOLANO 2014).
Las marcas abióticas, que se han podido analizar en esta Fase I, han sido el weathering, vermiculaciones y oxidaciones superficiales en el hueso (GRAYSON 1988; FISHER 1995; GIACOBONI Y PATOU-MATHIS 2002; BLASCO 2006; GARCÍA SOLANO 2014).
El weathering constituye exclusivamente el 3.8% de los restos, con 15 restos analizados con dicha alteración; mientras que las vermiculaciones o las marcas producidas por raíces presentan el 6.3% del conjunto, con 25 restos.
Las oxidaciones producidas por la acción hídrica se han podido constatar en solo 3 restos, representando solo el 0.7% del conjunto faunístico analizado de la Fase I.
Respecto a las alteraciones bióticas representadas en la Fase II, las marcas de corte no constituyen el principal proceso tafonómico de acción antrópica, en comparación con la Fase I. En su caso, constituyen el 0.5% de todos los restos analizados de dicha fase, con un total de 4 restos con marcas de corte, y el 28% de las marcas bióticas analizadas.
Dentro de estas marcas, se han podido distinguir tres tipos, entre las que se encuentran las marcas de desollado, evisceración y una marca indeterminada.
Las marcas de desollado, presentan el 50% de las marcas de corte de la Fase II, representadas en un cráneo y un hueso largo. La evisceración constituye exclusivamente el 25% restante, habiendo sido analizada gracias a una costilla; el resto con marca indeterminada corresponde a un hueso largo, y proporciona el 25% de las marcas de corte analizadas (Fig. 8).
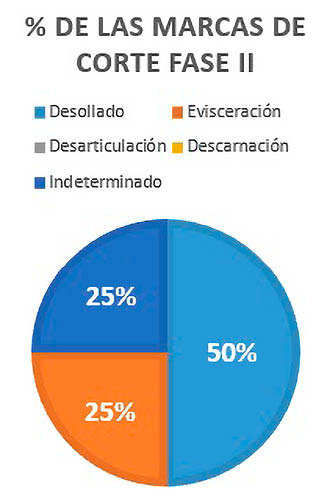
Fig. 8. Porcentajes de cada tipo de marca de corte analizada en la Fase II.
Las alteraciones térmicas, en la Fase II, presentan solo un 0.7% de todo el material analizado de dicha fase y el 43% de las marcas bióticas, con solo 6 restos quemados. Al igual que en la Fase I, las coloraciones tienden a ser grisáceas y blanquecinas en todo su conjunto, indicando de nuevo una intencionalidad de eliminación de estos residuos, y no una intención de cocinado.
Por último, las alteraciones producidas por carnívoros solo constituyen el 0.5% de todos los restos de la Fase I y el 29% de las marcas alteradas bióticamente, con 4 marcas o mordeduras analizadas.
Las marcas abióticas, que se han podido analizar en esta Fase II, han sido al igual que en la Fase I, el weathering, vermiculaciones y oxidaciones superficiales en el hueso.
El weathering constituye exclusivamente el 4.3% de los restos, es decir, 36 restos analizados con dicha alteración; mientras que las vermiculaciones o las marcas producidas por raíces presentan el 6.1% del conjunto, con 52 restos.
Las oxidaciones producidas por la acción del agua sobre el hueso se han podido constatar en 10 fragmentos, representando solo el 0.1% del conjunto faunístico analizado de la Fase II.
DISCUSIÓN Y CONCLUSIONES
El Polideportivo de Martos, en Jaén, constituye un yacimiento de gran importancia para el estudio y el conocimiento de las poblaciones del neolítico final en el Alto Guadalquivir. Las campañas de 1991 y 1993, nos han permitido acercarnos, aún más si cabe, al conocimiento de estas comunidades, dándonos una gran información socio-económica del momento.
La importancia y consideración de este presente trabajo radica en el exhaustivo y elaborado análisis llevado a cabo en 1.173 restos faunísticos del Complejo Estructural (CE) XV, confirmando una más que posible intencionalidad de limpieza y eliminación de residuos, retirados de las zonas de producción y consumo y sellados/enterrados en el complejo referida, dado el escaso número de marcas derivadas de procesos postdeposicionales e incluso de marcas de carnívoros. Tampoco las marcas antrópicas son abundantes en relación con la cantidad de restos lo que, sin duda, deriva en parte del carácter de deposición intencional de una gran parte de los huesos recuperados. (LIZCANO et al. 1991; CÁMARA et al. 2008, CÁMARA et al. 2010).
También se han podido corroborar los resultados taxonómicos obtenidos en estudios zooarqueológicos previos (LIZCANO et al. 1991; RIQUELME et al. 2012), y añadido a la lista de especies dos nuevos taxones de micromamíferos como son la rata (Rattus sp.) y el lirón careto (Eliomys Quercinus).
A pesar de que el conjunto faunístico estudiado y analizado del Complejo Estructural XV constituya una pequeña muestra de todo el conjunto óseo del Polideportivo de Martos, nos puede acercar a una mejor compresión de la economía que se pudo desarrollar en la zona durante el Neolítico Reciente.
Aunque nos encontremos con sociedades y agriculturas donde se podría guardar el excedente de cereal y leguminosas, el caso del Polideportivo de Martos, nos acerca, posiblemente, más a una sociedad ganadera que agricultora, ya que el material carpológico recuperado en este yacimiento, constituye una muy pequeña parte de lo que podría ser la alimentación de estas comunidades, a día de hoy.
Si bien el material faunístico es bastante más amplio, lo que nos daría como resultado una economía basada más en la ganadería, sobre todo por la gran cantidad de restos de animales domésticos en comparación con los animales salvajes.
Las especies domésticas que más protagonismo tendrían serían los animales pertenecientes al conjunto de los ovicaprinos (Capra hircus y Ovis aries) con un número mínimo de diez individuos, superando con gran diferencia a los animales de un mayor peso como son la vaca (Bos taurus) y el cerdo (Sus domesticus).
Aún con la complejidad para la determinación y la similitud entre algunas partes anatómicas de la oveja y la cabra, se han podido determinar generalmente un mayor número de restos de cabra (20 restos) que de oveja (6 restos), lo que nos podría indicar una mayor explotación de las cabras que de las ovejas, teniendo en cuenta que éstas son aparte de productoras de leche, también de lana.
La edad general para el consumo de los ovicaprinos sería una vez entrados en la edad adulta con siete restos, sin encontrar constancia de ovicápridos seniles, y solo dos individuos juveniles y un individuo infantil.
El ganado porcino sería la siguiente especie por detrás del ovicaprino en cuanto a importancia de explotación ganadera se refiere, con cinco individuos. Generalmente, se consumían antes de entrar en la fase adulta habiendo sido hallados mínimo dos individuos juveniles y uno infantil.
El ganado bovino, tendría una escasa representación en comparación con los dos ganados anteriores. Entre las dos fases, han aparecido tres individuos, los cuales dos pertenecen a ejemplares adultos (ocho restos determinados) y uno a un ternero ritual inhumado (75 restos determinados).
Fuera de la ganadería encontramos al perro como especie doméstica principal. En este caso, casi todos los restos pertenecen al perro inhumado en la Fase I, con 190 restos, correspondientes a un uso ritual. En el caso, de los 19 restos pertenecientes a la Fase II, y a un solo individuo, no pertenecerían, a priori, a un uso ritual como el anterior, aunque tampoco podemos aventurarnos a decir que tendría un uso de consumo, pues de los 19 restos de perro de la Fase I, no aparece ninguna marca antrópica en ellos.
Es posible, que en un futuro estudio de otros Complejos Estructurales del Polideportivo de Martos, se nos pueda arrojar un poco de luz sobre este tema.
Entre las especies salvajes más predominantes aparecen los lagomorfos con 39 restos y un mínimo de tres individuos entre el conejo (Oryctolagus cuniculus) y la liebre (Lepus granatensis).
El ciervo, por su parte, tendría una presencia de dos individuos entre las dos fases, con un total de 40 restos de los cuales 18 corresponden a fragmentos y restos de cornamenta.
Con un porcentaje mucho menos notable encontramos al caballo (Equus ferus) y al jabalí (Sus scrofa).
La presencia de caballo en el CE XV es bastante reducida con solo ocho restos y un mínimo de dos individuos; así como el jabalí cuya presencia se ha podido constatar gracias a dos piezas dentales.
Por lo tanto, el análisis arqueozoológico del conjunto faunístico del Complejo Estructural XV del Polideportivo de Martos analizado permite determinar una importancia ganadera en la economía de esta población, complementado con actividades cinegéticas, como la caza de lagomorfos principalmente.
A nivel tafonómico, se observa una muy escasa cantidad de huesos con marcas antrópicas y de carnívoros, aunque las marcas de corte que encontramos se ajustan a los datos expuestos anteriormente con los análisis anatómicos y taxonómicos, lo que nos indica que se consume más carne de origen ovicaprino con cuatro de los diez restos con marcas de corte halladas, cinco corresponderían a fragmentos de hueso largo indeterminados de animales pertenecientes a una talla media, y por último encontraríamos un resto con gran cantidad de marcas de corte en un fémur de équido.
En cuanto a las marcas de alteraciones térmicas, las coloraciones generales nos indican que la intención del uso del fuego podría ser la de limpiar y eliminar residuos, sin una intencionalidad de consumo basado en la cocina y cocción de carne.
Las marcas de carnívoros se han podido documentar solo en seis restos hallados, destacando sobre todo en los huesos del esqueleto axial como son las costillas, y en mayor medida de animales de gran talla, como serían: dos mordeduras en dos costillas diferentes de équido, y una mordedura en la costilla de un bóvido, aunque también han aparecido dos mordeduras en dos huesos largos.
Es importante remarcar que la pequeña representación de marcas antrópicas se pueda deber también a factores abióticos como podrían ser el weathering, las vermiculaciones o alteraciones producidas por acciones hídricas, entre otros, por lo que no es descartable que los porcentajes de alteraciones bióticas se puedan incrementar.
Por otra parte, la representatividad anatómica analizada nos permite conocer unos patrones de consumo más claros, donde las partes del esqueleto apendicular toman una mayor presencia, junto con las zonas de costillar de los ovicaprinos. Por lo tanto, cabe destacar que en los animales de mayor peso que los ovicaprinos destaca el consumo de las partes con mayor volumen de carne, mientras que en las especies de ovicápridos el aprovechamiento es igual de intenso en toda su anatomía.
Por lo tanto la importancia de los resultados plasmados en el presente trabajo, nos acercan a entender, aún más si cabe, estas poblaciones del Neolítico Reciente en la Alta Andalucía, dándonos una gran información socio-económica, confirmando nuevamente una más que probable intencionalidad de limpieza de los residuos en las zonas de producción y consumo dada la escasez de alteraciones bióticas en relación con la cantidad de restos óseos, depositados con carácter intencional, de individuos enterrados en conexión anatómica.
BIBLIOGRAFÍA
AFONSO, J.A; CÁMARA, J.A.; SPANEDDA, L.; ESQUIVEL, J.A.; LIZCANO, R.; PÉREZ, C. y RIQUELME, J.A. (2014): Nuevas aportaciones para la periodización del yacimiento del Polideportivo de Martos (Jaén): la evaluación estadística de las dataciones obtenidas para contextos rituales, Archivo de Prehistoria Levantina Vol. XXX, 2014, pp. 133-158.
CÁMARA. J.A.; LIZCAINO, R.; PÉREZ, C. y GÓMEZ, E. (2008): Apropiación, sacrificio, consumo y exhibición ritual de los animales en el Polideportivo de Martos. Sus implicaciones en los orígenes de la desigualdad social, Cuadernos de Prehistoria y Arqueología de la Universidad de Granada 18, 2008, pp. 55-90.
CÁMARA. J.A.; RIQUELME, J.A.; PÉREZ, C.; LIZCAINO, R.; BURGOS, A. y TORRERS, F. (2010): Sacrificio de animales y ritual en el Polideportivo de Martos-La Alberquilla (Martos, Jaén), Cuadernos de Prehistoria y Arqueología de la Universidad de Granada 20, 2010, pp. 295-328.
BINFORD, L.R. (1981): Bones, Ancient Men and Modern Myths, Academic Press Inc, 1981.
BLASCO, R. (2006): Estrategias de subsistencia de los homínidos del nivel XII de la Cova del Bolomor (La Valldigna, Valencia), Tesis de licenciatura Universitat Rovira i Virgili, Tarragona, 2006.
DOMÍNGUEZ-RODRIGO, M. y PIQUERAS, A. (2003): The use of tooth pits to identify carnivore taxa in tooth-marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours, Journal of Archaeological Science 30, 2003, pp. 1385-1391.
FERNÁNDEZ-JALVO, Y. y ANDRESW, P. (2016): Atlas of Taphonomic Identifications, Madrid. Ed. Springer, 2016.
FISHER, J. (1995): Bone Surface modifications in zooarchaeology, Journal of Archaeological Method and Theory 2(1), 1995, pp. 7-68.
GARCÍA SOLANO, J.A. (2014): La persistencia en las estrategias de subsistencia de los grupos humanos del Pleistoceno medio, a partir del registro fósil de la Cueva del Ángel (Lucena, Córdoba), Tesis doctoral Universidad de Granada. Granada, 2014.
GIACOBONI, G. y PATOU-MATHIS, M. (2002): Fiche rappels taphonomiques, Retouchoirs, compresseurs, percuteurs: os à impressions et éraillures, Société Préhistorique Française, Paris, 2002.
GRAYSON, D.K. (1988): Danger Cave, Last Supper Cave, and Haanging Rock Shelter: The Faunas, Anthropological Papers of American Museum of Natural History 66, 1988, pp. 1-130.
HEDGES, R.E.M. y MILLARD, A.R. (1995): Bones and Groundwater: Towards the Modelling of Diagenetic Processes, Journal of Archaeological Science 22(2), 1995, pp. 155-164.
LIRA GARRIDO, J.E. (2015) Rastreando los orígenes de la domesticación del caballo en Iberia: ADN antiguo y la evidencia de Atapuerca, Dentra Médica. Revista de Humanidades Vol. 14, 2, 2015, pp. 163-175.
LIRA GARRIDO, J.E. (2018) Estudio de la domesticación del caballo (Equus caballus) en la Península Ibérica a partir del análisis de ADN mitocondrial antiguo, Tesis Universidad Complutense de Madrid, 2018.
LIZCANO, R.; GÓMEZ, E.; CÁMARA, J.A.; AGUAYO, M.; ARAQUE, D.; BELLIDO, I.; CONTRERAS, L.; HERNÁNDEZ, M.; IZQUIERDO, M. y RUIZ, J. (1991): Primera campaña de excavación de urgencia en el Pabellón Polideportivo de Martos (Jaén), Anuario Arqueológico de Andalucía III, 1991, pp. 278-291.
RIQUELME, J.A.; LIZCANO, R.; PÉREZ, C.; SÁNCHEZ, R. y CÁMARA, J.A. (2012): Una introducción al análisis de los restos faunísticos de la zona arqueológica del Polideportivo de Martos, Cuadernos de Prehistoria y Arqueología de la Universidad de Granada 22, 2012, pp. 199-230.
SELVAGGIO, M.M. (1994): Carnivore tooth marks and stone tool butchery marks on scavenged bones: archaeological implications, Journal of Human Evolution 27, 1994, pp. 215-228.
